Фотосинтез на земле. Биологический процесс фотосинтеза и его значение в природе
Воду и минеральные вещества растения получают с помощью корней. Листья обеспечивают органическое питание растений. В отличие от корней они находятся не в почве, а в воздушной среде, поэтому осуществляют не почвенное, а воздушное питание.
Из истории изучения воздушного питания растений
Знания о питании растений накапливались постепенно. Около 350 лет назад голландский ученый Ян Гельмонт впервые поставил опыт по изучению питания растений. В глиняном горшке с почвой он выращивал иву, добавляя туда только воду. Опадавшие листья ученый тщательно взвешивал. Через пять лет масса ивы вместе с опавшими листьями увеличилась на 74,5 кг, а масса почвы уменьшилась всего на 57 г. На основании этого Гельмонт пришел к выводу, что все вещества в растении образуются не из почвы, а из воды. Мнение о том, что растение увеличивается в размерах только за счет воды, сохранялось до конца XVIII века.
В 1771 г. английский химик Джозеф Пристли изучал углекислый газ, или, как он его называл, «испорченный воздух» и сделал замечательное открытие. Если зажечь свечу и накрыть оо стеклянным колпаком, то, немного погорев, она погаснет. Мышь под таким колпаком начинает задыхаться. Однако если под колпак вместе с мышью поместить ветку мяты, то мышь не задыхается и продолжает жить. Значит, растения «исправляют» воздух, испорченный дыханием животных, то есть превращают углекислый газ в кислород.
В 1862 г. немецкий ботаник Юлиус Сакс с помощью опытов доказал, что зеленые растения не только выделяют кислород, но и создают органические вещества, служащие пищей всем другим организмам.
Фотосинтез
Главное отличие зеленых растений от других живых организмов - наличие в их клетках хлоропластов, содержащих хлорофилл. Хлорофилл обладает свойством улавливать солнечные лучи, энергия которых необходима для создания органических вещсств. Процесс образования органического вещества из углекислого газа и воды с помощью солнечной энергии называется фотосинтезом (греч. рЬо1оз свет). В процессе фотосинтеза образуются не только органические вещества - сахара, но и выделяется кислород.
Схематически процесс фотосинтеза можно изобразить так:
Вода поглощается корнями и по проводящей системе корней и стебля передвигается к листьям. Углекислый газ - составная часть воздуха. Он поступает в листья через открытые устьица. Поглощению углекислого газа способствует строение листа: плоская поверхность листовых пластинок, увеличивающая площадь соприкосновения с воздухом, и наличие большого числа устьиц в кожице.
Образующиеся в результате фотосинтеза сахара превращаются в крахмал. Крахмал это органическое вещество, которое не растворяется в воде. Кго легко обнаружить с помощью раствора йода.
Доказательства образования крахмала в листьях на свету
Докажем, что в зеленых листьях растений из углекислого газа и воды образуется крахмал. Для этого рассмотрим опыт, который в свое время был поставлен Юлиусом Саксом.
Комнатное растение (герань или примулу) выдерживают двое суток в темноте, чтобы весь крахмал израсходовался на процессы жизнедеятельности. Затем несколько листьев закрывают с двух сторон черной бумагой так, чтобы была прикрыта только их часть. Днем растение выставляют на свет, а ночью его дополнительно освещают с помощью настольной лампы.
Через сутки исследуемые листья срезают. Чтобы выяснить, в какой части листа образовался крахмал, листья кипятят в воле (чтобы набухли крахмальные зерна), а затем выдерживают в горячем спирте (хлорофилл при этом растворяется, и лист обесцвечивается). Затем листья промывают в воде и действуют на них слабым раствором йода. Тс участки листьев, которые были на свету, приобретают от действия йода синюю окраску. Это означает, что крахмал образовался в клетках освещенной части листа. Следовательно, фотосинтез происходит только на свету.

Доказательства необходимости углекислого газа для фотосинтеза
Чтобы доказать, что для образования крахмала в листьях необходим углекислый газ, комнатное растение также предварительно выдерживают в темноте. Затем один из листьев помещают в колбу с небольшим количеством известковой воды. Колбу закрывают ватным тампоном. Растение выставляют на свет. Углекислый газ поглощается известковой водой, поэтому его в колбе не будет. Лист срезается, и так же, как в предыдущем опыте, исследуется на наличие крахмала. Он выдерживается в горячей воде и спирте, обрабатывается раствором йода. Однако в этом случае результат опыта будет иным: лист не окрашивается в синий цвет, т.к. крахмал в нем не содержится. Следовательно, для образования крахмала, кроме света и воды, необходим углекислый газ.
Таким образом, мы ответили на вопрос, какую пищу получает растение из воздуха. Опыт показал, что это углекислый газ. Он необходим для образования органического вещества.
Организмы, самостоятельно создающие органические вещества для построения своего тела, называются автотрофамн (греч. autos - сам, trofe - пища).

Доказательства образования кислорода в процессе фотосинтеза
Чтобы доказать, что при фотосинтезе растения во внешнюю среду выделяют кислород, рассмотрим опыт с водным растением элодеей. Побеги элодеи опускают в сосуд с водой и сверху накрывают воронкой. На конец воронки надевают пробирку с водой. Растение выставляют на свет на двое-трое суток. На свету элодея выделяет пузырьки газа. Они скапливаются в верхней части пробирки, вытесняя воду. Для того чтобы выяснить, какой это газ, пробирку аккуратно снимают и вносят в нее тлеющую лучинку. Лучинка ярко вспыхивает. Это значит, что в колбе накопился газ, поддерживающий горение кислород.

Космическая роль растений
Растения, содержащие хлорофилл, способны усваивать солнечную энергию. Поэтому К.А. Тимирязев назвал их роль на Земле космической. Часть энергии Солнца, запасенная в органическом веществе, может долго сохраняться. Каменный уголь, торф, нефть образованы веществами, которые в далекие геологические времена были созданы зелеными растениями и вобрали в себя энергию Солнца. Сжигая природные горючие материалы, человек освобождает энергию, запасенную миллионы лет назад зелеными растениями.
Вопрос 1. Что такое фотосинтез? Назовите вещества, необходимые для его осуществления.
Фотосинтез – это процесс образования органических веществ и кислорода из углекислого газа и воды в листьях зеленых растений на солнечном свету.
Вопрос 2. Закончите предложения.
Фотосинтез происходит в растительных клетках, которые содержат органоиды хлоропласты. В них содержится зелёный пигмент хлорофилл, который придает растению окраску и обеспечивает фотосинтез.
У большинства растений основным органом, обеспечивающим осуществление фотосинтеза, является лист, еще фотосинтез может протекать в стеблях и зеленых плодах.
Вопрос 3. Известно, что наземные растения ежегодно образуют столько листьев, что ими можно было бы покрыть земной шар в несколько слоёв. Объясните, почему у растений образуется так много листьев.
Процесс образования органических веществ идет в листьях зеленых астений на солнечном свету. Поэтому, чтобы прокормить растение листьев должно быть очень много.
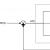
Вопрос 4. Рассмотрите рисунок «Образование органических веществ в процессе фотосинтеза». Подпишите на нем названия веществ, поступающих в лист и выводящихся из него.
Углекислый газ
Кислород
Ответьте на вопросы:
1) Каковы необходимые условия осуществления фотосинтеза?
Для фотосинтеза необходим солнечный свет, углекислый газ и хлоропласты.
2) Какие органические вещества образуются в процессе фотосинтеза и каково их значение для растения?
В хлоропластах под воздействием света в процессе фотосинтеза у растений образуется крахмал. Это вещество является углеводом и служит источником энергии для растений.
Вопрос 5*. Прочитайте в учебнике описание опыта по изучению влияния света на образование органических веществ в зеленых растениях и рассмотрите рисунок 61. Как вы думаете, почему в листьях зеленых растений нельзя обнаружить крахмал, после того как их выдерживают в темноте в течение 2-3 дней? Куда он исчезает?
Для преобразования крахмала в листьях необходим солнечный свет. Крахмал образуется в процессе фотосинтеза. Этот процесс произойдет с использованием энергии света. Без света нет процесса фотосинтеза, без процесса нет в листьях крахмала.
Работаем в лаборатории
Вопрос 6. Рассмотрите рисунок, на котором изображен опыт.
Ответьте на вопросы:
1) Почему свеча в первом и третьем случаях гаснет?
В первом и третьем сосудах семена и корнеплоды в процессе дыхания истратили весь кислород и выделили углекислый газ. Свеча погасла.
2) Почему свеча во втором случае горит?
Во втором сосуде растение не только дышит, но и при помощи фотосинтеза выделяет кислород, поэтому свеча горит.
Процесс преобразования лучистой энергии Солнца в химическую с использованием последней в синтезе углеводов из углекислого газа. Это единственный путь улавливания солнечной энергии и использования ее для жизни на нашей планете.
Улавливание и преобразование солнечной энергии осуществляют многообразные фотосинтезирующие организмы (фотоавтотрофы). К ним относятся многоклеточные организмы (высшие зеленые растения и низшие их формы - зеленые, бурые и красные водоросли) и одноклеточные (эвгленовые, динофлагелляты и диатомовые водоросли). Большую группу фотосинтезирующих организмов составляют прокариоты - сине-зеленые водоросли, зеленые и пурпурные бактерии. Примерно половина работы по фотосинтезу на Земле осуществляется высшими зелеными растениями, а остальная половина - главным образом одноклеточными водорослями.
Первые представления о фотосинтезе были сформированы в 17 веке. В дальнейшем, по мере появления новых данных, эти представления многократно изменялись [показать] .
Развитие представлений о фотосинтезе
Начало изучению фотосинтеза было положено в 1630 году, когда ван Гельмонт показал, что растения сами образуют органические вещества, а не получают их из почвы. Взвешивая горшок с землей, в котором росла ива, и само дерево, он показал, что в течение 5 лет масса дерева увеличилась на 74 кг, тогда как почва потеряла только 57 г. Ван Гельмонт пришел к заключению, что остальную часть пищи растение получило из воды, которой поливали дерево. Теперь мы знаем, что основным материалом для синтеза служит двуокись углерода, извлекаемая растением из воздуха.
В 1772 году Джозеф Пристли показал, что побег мяты "исправляет" воздух, "испорченный" горящей свечой. Семь лет спустя Ян Ингенхуз обнаружил, что растения могут "исправлять" плохой воздух только находясь на свету, причем способность растений "исправлять" воздух пропорциональна ясности дня и длительности пребывания растений на солнце. В темноте же растения выделяют воздух, "вредный для животных".
Следующей важной ступенью в развитии знаний о фотосинтезе были опыты Соссюра, проведенные в 1804 году. Взвешивая воздух и растения до фотосинтеза и после, Соссюр установил, что увеличение сухой массы растения превышало массу поглощенной им из воздуха углекислоты. Соссюр пришел к выводу, что другим веществом, участвовавшим в увеличении массы, была вода. Таким образом, 160 лет назад процесс фотосинтеза представляли себе следующим образом:
H 2 O + CO 2 + hv -> C 6 H 12 O 6 + O 2
Вода + Углекислота + Солнечная энергия ----> Органическое вещество + Кислород
Ингенхуз предположил, что роль света в фотосинтезе заключается в расщеплении углекислоты; при этом происходит выделение кислорода, а освободившийся "углерод" используется для построения растительных тканей. На этом основании живые организмы были разделены на зеленые растения, которые могут использовать солнечную энергию для "ассимиляции" углекислоты, и остальные организмы, не содержащие хлорофилла, которые не могут использовать энергию света и не способны ассимилировать CO 2 .
Этот принцип разделения живого мира был нарушен, когда С. Н. Виноградский в 1887 году открыл хемосинтезирующие бактерии - бесхлорофильные организмы, способные ассимилировать (т. е. превращать в органические соединения) углекислоту в темноте. Он был нарушен также, когда в 1883 году Энгельман открыл пурпурные бактерии, осуществляющие своеобразный фотосинтез, не сопровождающийся выделением кислорода. В свое время этот факт не был оценен в должной мере; между тем открытие хемосинтезирующих бактерий, ассимилирующих углекислоту в темноте, показывает, что ассимиляцию углекислоты нельзя считать специфической особенностью одного лишь фотосинтеза.
После 1940 года благодаря применению меченого углерода было установлено, что все клетки - растительные, бактериальные и животные - способны ассимилировать углекислоту, т. е. включать ее в состав молекул органических веществ; различны лишь источники, из которых они черпают необходимую для этого энергию.
Другой крупный вклад в изучение процесса фотосинтеза внес в 1905 году Блэкман, который обнаружил, что фотосинтез состоит из двух последовательных реакций: быстрой световой реакции и ряда более медленных, не зависящих от света этапов, названных им темповой реакцией. Используя свет высокой интенсивности, Блэкман показал, что фотосинтез протекает с одинаковой скоростью как при прерывистом освещении с продолжительностью вспышек всего в долю секунды, так и при непрерывном освещении, несмотря на то что в первом случае фотосинтетическая система получает вдвое меньше энергии. Интенсивность фотосинтеза снижалась только при значительном увеличении темнового периода. В дальнейших исследованиях было установлено, что скорость темновой реакции значительно возрастает с повышением температуры.
Следующая гипотеза относительно химической основы фотосинтеза была выдвинута ван Нилем, который в 1931 году экспериментально показал, что у бактерий фотосинтез может происходить в анаэробных условиях, не сопровождаясь выделением кислорода. Ван Ниль высказал предположение, что в принципе процесс фотосинтеза сходен у бактерий и у зеленых растений. У последних световая энергия используется для фотолиза воды (Н 2 0) с образованием восстановителя (Н), определенным путем участвующего в ассимиляции углекислоты, и окислителя (ОН) - гипотетического предшественника молекулярного кислорода. У бактерий фотосинтез протекает в общем так же, но донором водорода служит Н 2 S или молекулярный водород, и поэтому выделения кислорода не происходит.
Современные представления о фотосинтезе
По современным представлениям сущность фотосинтеза заключается в превращении лучистой энергии солнечного света в химическую энергию в форме АТФ и восстановленного никотинамидадениндинуклеотидфосфата (НАДФ · Н).
В настоящее время принято считать, что процесс фотосинтеза складывается из двух стадий, в которых активное участие принимают фотосинтезирующие структуры [показать] и светочувствительные пигменты клетки .
Фотосинтезирующие структуры
У бактерий фотосинтезирующие структуры представлены в виде впячивания клеточной мембраны, образуя пластинчатые органоиды мезосомы. Изолированные мезосомы, получаемые при разрушении бактерий, называются хроматофорами, в них сосредоточен светочувствительный аппарат.
У эукариотов фотосинтетический аппарат расположен в специальных внутриклеточных органоидах - хлоропластах, содержащих зеленый пигмент хлорофилл, который придает растению зеленую окраску и играет важнейшую роль в фотосинтезе, улавливая энергию солнечного света. Хлоропласты, подобно митохондриям, содержат также ДНК, РНК и аппарат для синтеза белка, т. е. обладают потенциальной способностью к самовоспроизведению. По размерам хлоропласты в несколько раз больше митохондрий. Число хлоропластов колеблется от одного у водорослей до 40 на клетку у высших растений.

В клетках зеленых растений помимо хлоропластов имеются и митохондрии, которые используются для образования энергии в ночное время за счет дыхания, как в гетеротрофных клетках.
Хлоропласты имеют шаровидную или уплощенную форму. Они окружены двумя мембранами - наружной и внутренней (рис. 1). Внутренняя мембрана укладывается в виде стопок уплощенных пузырьковидных дисков. Эта стопка называется граной.
Каждая грана состоит из отдельных слоев, расположенных наподобие столбиков монет. Слои белковых молекул чередуются со слоями, содержащими хлорофилл, каротины и другие пигменты, а также особые формы липидов (содержащих галактозу или серу, но только одну жирную кислоту). Эти поверхностно-активные липиды, по-видимому, адсорбированы между отдельными слоями молекул и служат для стабилизации структуры, состоящей из чередующихся слоев белка и пигментов. Такое слоистое (ламеллярное) строение граны, вероятнее всего облегчает перенос энергии в процессе фотосинтеза от одной молекулы к близлежащей.
В водорослях находится не более одной граны в каждом хлоропласте, а в высших растениях - до 50 гран, которые соединены между собой мембранными перемычками. Водная среда между гранами - это строма хлоропласта, которая содержит ферменты, осуществляющие "темновые реакции"
Пузырьковидные структуры, из которых состоит грана, называются тилактоидами. В гране от 10 до 20 тилактоидов.
Элементарная структурная и функциональная единица фотосинтеза мембран тилактоидов, содержащая необходимые светоулавливающие пигменты и и компоненты аппарата трансформации энергии, называется квантосомой, состоящей примерно из 230 молекул хлорофилла. Эта частица имеет массу порядка 2 х 10 6 дальтон и размеры около 17,5 нм.
Стадии фотосинтеза |
||
Световая стадия (или энергетическая) |
Темновая стадия (или метаболическая) |
|
Место протекание реакции |
В квантосомах мембран тилактоидов, протекает на свету. |
Осуществляется вне тилактоидов, в водной среде стромы. |
Начальные продукты |
Энергия света, вода (Н 2 О), АДФ, хлорофилл |
СО 2 , рибулозодифосфат, АТФ, НАДФН 2 |
Суть процесса |
Фотолиз воды, фосфорилирование В световой стадии фотосинтеза энергия света трансформируется в химическую энергию АТФ, а бедные энергией электроны воды переходят в богатые энергией электроны НАДФ· Н 2 . Побочным веществом, образующимся в ходе световой стадии, является кислород. Реакции световой стадии получили название "световых реакций". |
Карбоксилирование, гидрирование, дефосфорилирование В темновой стадии фотосинтеза протекают "темновые реакции" при которых наблюдается восстановительный синтез глюкозы из CO 2 . Без энергии световой стадии темновая стадия невозможна. |
Конечные продукты |
О 2 , АТФ, НАДФН 2 Богатые энергией продукты световой реакции - АТФ и НАДФ· Н 2 далее используются в темновой стадии фотосинтеза. |
|
Взаимосвязь между световой и темновой стадиями можно выразить схемой
Процесс фотосинтеза эндергонический, т.е. сопровождается увеличением свободной энергии, поэтому требует значительного количества энергии, подведенной извне. Суммарное уравнение фотосинтеза: 6СО 2 + 12Н 2 О--->С 6 Н 12 О 62 + 6Н 2 О + 6О 2 + 2861 кДж/моль. |
||

Наземные растения поглощают необходимую для процесса фотосинтеза воду через корни, а водные растения получают ее путем диффузии из окружающей среды. Необходимая для фотосинтеза углекислота диффундирует в растение через мелкие отверстия на поверхности листьев - устьица. Поскольку углекислота расходуется в процессе фотосинтеза, ее концентрация в клетке обычно несколько ниже, чем в атмосфере. Освобождающийся в процессе фотосинтеза кислород диффундирует наружу из клетки, а затем и из растения - через устьица. Образующиеся при фотосинтезе сахара также диффундируют в те части растения, где их концентрация ниже.
Для осуществления фотосинтеза растениям необходимо очень много воздуха, так как он содержит всего 0,03% углекислоты. Следовательно, из 10 000 м 3 воздуха можно получить 3 м 3 углекислоты, из которой в процессе фотосинтеза образуется около 110 г глюкозы. Обычно растения лучше растут при более высоком содержании в воздухе углекислоты. Поэтому в некоторых теплицах содержание CO 2 в воздухе доводят до 1-5%.
Механизм световой (фотохимической) стадии фотосинтеза |
В реализации фотохимической функции фотосинтеза принимают участие солнечная энергия и различные пигменты: зеленые - хлорофиллы а и b, желтые - каротиноиды и красные или синие - фикобилины. Фотохимически активен среди этого комплекса пигментов только хлорофилл а. Остальные пигменты играют вспомогательную роль, являясь лишь собирателями световых квантов (своеобразные светособирающие линзы) и проводниками их к фотохимическому центру.
На основании способности хлорофилла эффективно поглощать солнечную энергию определенной длины волны в мембранах тилактоидов были выделены функциональные фотохимические центры или фотосистемы (рис. 3):

- фотосистемa I (хлорофилл а ) - содержит пигмент 700 (Р 700) поглощающий свет с длиной волны около 700 нм, играет основную роль в образовании продуктов световой стадии фотосинтеза: АТФ и НАДФ · Н 2
- фотосистема II (хлорофилл b ) - содержит пигмент 680 (Р 680), поглощающий свет с длиной волны 680 нм, играет вспомогательную роль восполняя за счет фотолиза воды утраченные фотосистемой I электроны
На 300-400 молекул светособирающих пигментов в фотосистемах I и II приходится только одна молекула фотохимически активного пигмента - хлорофилла а.
Поглощенный растением световой квант
- переводит пигмент Р 700 из основного состояния в возбужденное - Р * 700 , в котором он легко теряет электрон с образованием
положительной электронной дырки в виде Р 700 + по схеме:
Р 700 ---> Р * 700 ---> Р + 700 + е -
После чего молекула пигмента, потерявшая электрон, может служить акцептором электрона (способна принять электрон) и переходить в восстановленную форму
- вызывает разложение (фотоокисление) воды в фотохимическом центре Р 680 фотосистемы II по схеме
Н 2 О ---> 2Н + + 2е - + 1/2O 2
Фотолиз воды называется реакцией Хилла. Электроны, образующиеся при разложении воды, первоначально акцептируются веществом, обозначаемым Q (иногда его называют цитохромом С 550 пo максимуму поглощения, хотя оно не является цитохромом). Затем от вещества Q через цепь переносчиков, похожую по составу на митохондриальную, электроны поставляются в фотосистему I для заполнения электронной дырки, образовавшейся в результате поглощения системой световых квантов, и восстановления пигмента Р + 700
Если такая молекула просто получит назад тот же электрон, то произойдет выделение световой энергии в виде тепла и флуоресценции (этим обусловлена флуоресценция чистого хлорофилла). Однако, в большинстве случаев, освободившийся отрицательно заряженный электрон акцептируется специальными железосерными белками (FеS-центр), а затем
- или транспортируется по одной из цепей переносчиков обратно к Р + 700 , заполняя электронную дырку
- или по другой цепи переносчиков через ферредоксин и флавопротеид к постоянному акцептору - НАДФ · Н 2
В первом случае происходит замкнутый циклический транспорт электрона, а во втором - нециклический.
Оба процесса катализируются одной и той же цепью переносчиков электронов. Однако при циклическом фотофосфорилировании электроны возвращаются от хлорофилла а снова к хлорофиллу а , тогда как при нециклическом фотофосфорилировании электроны переходят от хлорофилла b к хлорофиллу а .
| Циклическое (фотосинтетическое) фосфорилирование | Нециклическое фосфорилирование |
В результате циклического фосфорилирования происходит образование молекул АТФ. Процесс связан с возвращением через ряд последовательных этапов возбужденных электронов на Р 700 . Возвращение возбужденных электронов на Р 700 приводит к высвобождению энергии (при переходе с высокого на низкий энергетический уровень), которая, при участии фосфорилирующей ферментной системы, аккумулируется в фосфатных связях АТФ, а не рассеивается в виде флуоресценции и тепла (рис.4.). Этот процесс называется фотосинтетическим фосфорилированием (в отличие от окислительного фосфорилирования, осуществляемого митохондриями); Фотосинтетическое фосфорилирование - первичная реакция фотосинтеза - механизм образования химической энергии (синтеза АТФ из АДФ и неорганического фосфата) на мембране тилактоидов хлоропластов с использованием энергии солнечного света. Необходима для темновой реакции ассимиляции СО 2 |
В результате нециклического фосфорилирования происходит восстановление НАДФ + с образование НАДФ · Н. Процесс связан с передачей электрона ферредоксину, его восстановлением и дальнейшим переходом его к НАДФ + с последующим восстановление его до НАДФ · Н |
В тилактоидах идут оба процесса, хотя второй более сложный. Он сопряжен (взаимосвязан) с работой фотосистемы II. |
|


Таким образом, утраченные Р 700 электроны восполняются за счет электронов воды, разлагаемой под действием света в фотосистеме II.
а + в основное состояние, образуются, по-видимому, при возбуждении хлорофилла b . Эти высокоэнергетические электроны переходят к ферредоксину и затем через флавопротеин и цитохромы - к хлорофиллу а . На последнем этапе происходит фосфорилирование АДФ до АТФ (рис. 5).
Электроны, необходимые для возвращения хлорофилла в его основное состояние, поставляются, вероятно, ионами ОН - , образующимися при диссоциации воды. Некоторая часть молекул воды диссоциирует на ионы Н + и ОН - . В результате потери электронов ионы ОН - превращаются в радикалы (ОН), которые в дальнейшем дают молекулы воды и газообразного кислорода (рис. 6).
Этот аспект теории подтверждается результатами опытов с водой и CO 2 , меченными 18 0 [показать] .

Согласно этим результатам, весь газообразный кислород, выделяющийся при фотосинтезе, происходит из воды, а не из СО 2 . Реакции расщепления воды до сих пор еще подробно не изучены. Ясно, однако, что осуществление всех последовательных реакций нециклического фотофосфорилирования (рис. 5), в том числе возбуждение одной молекулы хлорофилла а и одной молекулы хлорофилла b , должно приводить к образованию одной молекулы НАДФ · Н, двух или более молекул АТФ из АДФ и Ф н и к выделению одного атома кислорода. Для этого необходимо по крайней мере четыре кванта света - по два для каждой молекулы хлорофилла.
Нециклический поток электронов от Н 2 О к НАДФ · Н 2 , происходящий при взаимодействии двух фотосистем и связывающих их электронно-транспортных цепей, наблюдается вопреки значениям редокс-потенциалов: Е° для 1/2O 2 /Н 2 О = +0,81 В, а Е° для НАДФ/НАДФ · Н = -0,32 В. Энергия света обращает поток электронов "вспять". Существенно то, что при переносе от фотосистемы II к фотосистеме I часть энергии электронов аккумулируется в виде протонного потенциала на мембране тилактоидов, а затем в энергию АТФ.

Механизм образования протонного потенциала в цепи переноса электронов и его использование на образование АТФ в хлоропластах сходен с таковым в митохондриях. Однако в механизме фотофосфорилирования имеются некоторые особенности. Тилактоиды представляют собой как бы вывернутые наизнанку митохондрии, поэтому направление переноса электронов и протонов через мембрану противоположно направлению его в митохондриальной мембране (рис.6). Электроны движутся к внешней стороне, а протоны концентрируются внутри тилактоидного матрикса. Матрикс заряжается положительно, а внешняя мембрана тилактоида - отрицательно, т. е. направление протонного градиента противоположно направлению его в митохондриях.
Другой особенностью является значительно большая доля рН в протонном потенциале по сравнению с митохондриями. Тилактоидный матрикс сильно закисляется, поэтому Δ рН может достигать 0,1-0,2 В, в то время как Δ Ψ составляет около 0,1 В. Общее значение Δ μ H+ > 0,25 В.
Н + -АТФ-синтетаза, обозначаемая в хлоропластах как комплекс "СF 1 +F 0 ", ориентирована тоже в противоположном направлении. Головка ее (F 1) смотрит наружу, в сторону стромы хлоропласта. Протоны выталкиваются через СF 0 +F 1 из матрикса наружу, и в активном центре F 1 образуется АТФ за счет энергии протонного потенциала.
В отличие от митохондриальной цепи в тилактоидной имеется, по-видимому, только два участка сопряжения, поэтому на синтез одной молекулы АТФ требуется вместо двух три протона, т. е. соотношение 3 Н + /1 моль АТФ.
Итак, на первой стадии фотосинтеза, во время световых реакций, в строме хлоропласта образуются АТФ и НАДФ · Н - продукты, необходимые для осуществления темновых реакций.


Механизм темновой стадии фотосинтеза |
Темновые реакции фотосинтеза - это процесс включения углекислоты в органические вещества с образованием углеводов (фотосинтез глюкозы из СО 2). Реакции протекают в строме хлоропласта при участии продуктов световой стадии фотосинтеза - АТФ и НАДФ · Н2.
Ассимиляция диоксида углерода (фотохимическое карбоксилирование) представляет собой циклический процесс, который называется также пентозофосфатным фотосинтетическим циклом или циклом Кальвина (рис. 7). В нем можно выделить три основные фазы:
- карбоксилирование (фиксация СО 2 рибулозодифосфатом)
- восстановление (образование триозофосфатов при восстановлении 3-фосфоглицерата)
- регенерация рибулозодифосфата
Рибулозо-5-фосфат (сахар, содержащий 5 атомов углерода, с фосфатным остатком у углерода в положении 5) подвергается фосфорилированию за счет АТФ, что приводит к образованию рибулозодифосфата. Это последнее вещество карбоксилируется путем присоединения СО 2 , по-видимому, до промежуточного шестиуглеродного продукта, который, однако, немедленно расщепляется с присоединением молекулы воды, образуя две молекулы фосфоглицериновой кислоты. Затем фосфоглицериновая кислота восстанавливается в ходе ферментативной реакции, для осуществления которой необходимо присутствие АТФ и НАДФ · Н с образованием фосфоглицеринового альдегида (трехуглеродный сахар - триоза). В результате конденсации двух таких триоз образуется молекула гексозы, которая может включаться в молекулу крахмала и таким образом откладываться про запас.
Для завершения этой фазы цикла в процессе фотосинтеза поглощается 1 молекула С0 2 и используются 3 молекулы АТФ и 4 атома Н (присоединенных к 2 молекулам НАД · Н). Из гексозофосфата путем определенных реакций пентозофосфатного цикла (рис. 8) регенерирует рибулозофосфат, который снова может присоединить к себе другую молекулу углекислоты.
Ни одну из описанных реакций - карбоксилирование, восстановление или регенерацию - нельзя считать специфичной только для фотосинтезирующей клетки. Единственное обнаруженное у них отличие заключается в том, что для реакции восстановления, в течение которой фосфоглицериновая кислота превращается в фосфоглицериновый альдегид, необходим НАДФ · Н, а не НАД · Н, как обычно.
Фиксация СО 2 рибулозодифосфатом катализируется ферментом рибулозодифосфаткарбоксилазой: Рибулозодифосфат + СО 2 --> 3-Фосфоглицерат Далее 3-фосфоглицерат восстанавливается с помощью НАДФ · Н 2 и АТФ до глицеральдегид-3-фосфата. Эта реакция катализируется ферментом - глицеральдегид-3-фосфат-дегидрогеназой. Глицеральдегид-3-фосфат легко изомеризуется в дигидроксиацетонфосфат. Оба триозофосфата используются в образовании фруктозобисфосфата (обратная реакция, катализируемая фруктозо-бисфосфат-альдолазой). Часть молекул образовавшегося фруктозобисфосфата участвует вместе с триозофосфатами в регенерации рибулозодифосфата (замыкают цикл), а другая часть используется для запасания углеводов в фотосинтезирующих клетках, как показано на схеме.
Подсчитано, что для синтеза одной молекулы глюкозы из СО 2 в цикле Кальвина требуется 12 НАДФ · Н + Н + и 18 АТФ (12 молекул АТФ расходуются на восстановление 3-фосфоглицерата, а 6 молекул - в реакциях регенерации рибулозодифосфата). Минимальное соотношение - 3 АТФ: 2 НАДФ · Н 2 .
Можно заметить общность принципов, лежащих в основе фотосинтетического и окислительного фосфорилирования, причем фотофосфорилирование представляет собой как бы обращенное окислительное фосфорилирование:

Энергия света является движущей силой фосфорилирования и синтеза органических веществ (S-Н 2) при фотосинтезе и, наоборот, энергия окисления органических веществ - при окислительном фосфорилировании. Поэтому именно растения обеспечивают жизнь животным и другим гетеротрофным организмам:

Углеводы, образующиеся при фотосинтезе, служат для построения углеродных скелетов многочисленных органических веществ растений. Азоторганические вещества усваиваются фотосинтезирующими организмами путем восстановления неорганических нитратов или атмосферного азота, а сера - восстановлением сульфатов до сульфгидрильных групп аминокислот. Фотосинтез в конечном итоге обеспечивает построение не только обязательных для жизни белков, нуклеиновых кислот, углеводов, липидов, кофакторов, но и многочисленных продуктов вторичного синтеза, являющихся ценными лекарственными веществами (алкалоиды, флавоноиды, полифенолы, терпены, стероиды, органические кислоты и т.д.).
Бесхлорофильный фотосинтез
Бесхлорофильный фотосинтез обнаружен у солелюбивых бактерий, имеющих фиолетовый светочувствительный пигмент. Этим пигментом оказался белок бактериородопсин, содержащий, подобно зрительному пурпуру сетчатки - родопсину, производное витамина А - ретиналь. Бактериородопсин, встроенный в мембрану солелюбивных бактерий, образует на этой мембране в ответ на поглощение ретиналем света протонный потенциал, преобразующийся в АТФ. Таким образом, бактериородопсин является бесхлорофильным преобразователем энергии света.
Фотосинтез и внешняя среда
Фотосинтез возможен только при наличии света, воды и диоксида углерода. КПД фотосинтеза не более 20% у культурных видов растений, а обычно он не превышает 6-7%. В атмосфере примерно 0,03% (об.) СО 2 , при повышении его содержания до 0,1% интенсивность фотосинтеза и продуктивность растений возрастают, поэтому целесообразно подкармливать растения гидрокарбонатами. Однако содержание СО 2 в воздухе выше 1,0% оказывает вредное действие на фотосинтез. За год только наземные растения усваивают 3% всего СО 2 атмосферы Земли, т. е. около 20 млрд. т. В составе синтезируемых из СО 2 углеводов аккумулируется до 4 · 10 18 кДж энергии света. Это соответствует мощности электростанции в 40 млрд кВт. Побочный продукт фотосинтеза - кислород - жизненно необходим для высших организмов и аэробных микроорганизмов. Сохранить растительный покров - значит сохранить жизнь на Земле.

Эффективность фотосинтеза
Эффективность фотосинтеза с точки зрения производства биомассы можно оценить через долю общей солнечной радиации, попадающей на определенную площадь за определенное время, которая запасается в органических веществах урожая. Продуктивность системы можно оценить по количеству органического сухого вещества, получаемого с единицы площади за год, и выразить в единицах массы (кг) или энергии (мДж) продукции, полученной с гектара за год.
Выход биомассы зависит, таким образом, от площади коллектора солнечной энергии (листьев), функционирующих в течение года, и числа дней в году с такими условиями освещенности, когда возможен фотосинтез с максимальной скоростью, что определяет эффективность всего процесса. Результаты определения доли солнечной радиации (в %), доступной растениям (фотосинтетически активной радиации, ФАР), и знание основных фотохимических и биохимических процессов и их термодинамической, эффективности позволяют рассчитать вероятные предельные скорости образования органических веществ в пересчете на углеводы.
Растения используют свет с длиной волны от 400 до 700 нм, т. е. на долю фотосинтетически активной радиации приходится 50% всего солнечного света. Это соответствует интенсивности на поверхности Земли 800-1000 Вт/м 2 за обычный солнечный день (в среднем). Усредненная максимальная эффективность превращения энергии при фотосинтезе на практике составляет 5-6%. Эти оценки получены на основе изучения процесса связывания СО 2 , а также сопутствующих физиологических и физических потерь. Одному молю связанного СО 2 в форме углевода соответствует энергия 0,47 МДж, а энергия моля квантов красного света с длиной волны 680 нм (наиболее бедный энергией свет, используемый в фотосинтезе) составляет 0,176 МДж. Таким образом, минимальное число молей квантов красного света, необходимое для связывания 1 моля СО 2 , составляет 0,47:0,176 = 2,7. Однако, поскольку перенос четырех электронов от воды для фиксации одной молекулы СО 2 требует не менее восьми квантов света, теоретическая эффективность связывания равна 2,7:8 = 33%. Эти расчеты сделаны для красного света; ясно, что для белого света эта величина будет соответственно ниже.
В наилучших полевых условиях эффективность фиксации в растениях достигает 3%, однако это возможно лишь в короткие периоды роста и, если пересчитать ее на весь год, то она будет где-то между 1 и 3%.
На практике в среднем за год эффективность фотосинтетического преобразования энергии в зонах с умеренным климатом составляет обычно 0,5-1,3%, а для субтропических культур - 0,5-2,5%. Выход продукта, который можно ожидать при определенном уровне интенсивности солнечного света и разной эффективности фотосинтеза, легко оценить из графиков, приведенных на рис. 9.
Значение фотосинтеза
- Процесс фотосинтеза является основой питания всех живых существ, а также снабжает человечество топливом, волокнами и бесчисленными полезными химическими соединениями.
- Из диоксида углерода и воды, связанных из воздуха в ходе фотосинтеза, образуется около 90-95% сухого веса урожая.
- Человек использует около 7% продуктов фотосинтеза в пищу, в качестве корма для животных, в виде топлива и строительных материалов
ФОТОСИНТЕЗ
образование живыми растительными клетками органических веществ, таких, как сахара и крахмал, из неорганических - из СО2 и воды - с помощью энергии света, поглощаемого пигментами растений. Это процесс производства пищи, от которого зависят все живые существа - растения, животные и человек. У всех наземных растений и у большей части водных в ходе фотосинтеза выделяется кислород. Некоторым организмам, однако, свойственны другие виды фотосинтеза, проходящие без выделения кислорода. Главную реакцию фотосинтеза, идущего с выделением кислорода, можно записать в следующем виде:
К органическим веществам относятся все соединения углерода за исключением его оксидов и нитридов. В наибольшем количестве образуются при фотосинтезе такие органические вещества, как углеводы (в первую очередь сахара и крахмал), аминокислоты (из которых строятся белки) и, наконец, жирные кислоты (которые в сочетании с глицерофосфатом служат материалом для синтеза жиров). Из неорганических веществ для синтеза всех этих соединений требуются вода (Н2О) и диоксид углерода (СО2). Для аминокислот требуются, кроме того, азот и сера. Растения могут поглощать эти элементы в форме их оксидов, нитрата (NO3-) и сульфата (SO42-) или в других, более восстановленных формах, таких, как аммиак (NH3) или сероводород (сульфид водорода H2S). В состав органических соединений может включаться при фотосинтезе также фосфор (растения поглощают его в виде фосфата) и ионы металлов - железа и магния. Марганец и некоторые другие элементы тоже необходимы для фотосинтеза, но лишь в следовых количествах. У наземных растений все эти неорганические соединения, за исключением СО2, поступают через корни. СО2 растения получают из атмосферного воздуха, в котором средняя его концентрация составляет 0,03%. СО2 поступает в листья, а О2 выделяется из них через небольшие отверстия в эпидермисе, называемые устьицами. Открывание и закрывание устьиц регулируют особые клетки - их называют замыкающими - тоже зеленые и способные осуществлять фотосинтез.
Когда на замыкающие клетки падает свет, в них начинается фотосинтез. Накопление его продуктов вынуждает эти клетки растягиваться. При этом устьичное отверстие открывается шире, и СО2 проникает к нижележащим слоям листа, клетки которых могут теперь продолжать фотосинтез. Устьица регулируют и испарение воды листьями, т.н. транспирацию, поскольку большая часть водяных паров проходит именно через эти отверстия. Водные растения добывают все необходимые им питательные вещества из воды, в которой живут. СО2 и ион бикарбоната (HCO3-) тоже содержатся и в морской, и в пресной воде. Водоросли и другие водные растения получают их непосредственно из воды. Свет в фотосинтезе играет роль не только катализатора, но и одного из реагентов. Значительная часть световой энергии, используемой растениями при фотосинтезе, запасается в виде химической потенциальной энергии в продуктах фотосинтеза. Для фотосинтеза, идущего с выделением кислорода, в той или иной мере пригоден любой видимый свет от фиолетового (длина волны 400 нм) до среднего красного (700 нм). При некоторых видах бактериального фотосинтеза, не сопровождающегося выделением O2, может эффективно использоваться свет с большей длиной волны, вплоть до дальнего красного (900 нм). Выяснение природы фотосинтеза началось еще во времена зарождения современной химии. Работы Дж.Пристли (1772), Я.Ингенхауза (1780), Ж.Сенебье (1782), а также химические исследования А.Лавуазье (1775, 1781) позволили сделать вывод, что растения превращают диоксид углерода в кислород и для этого процесса необходим свет. Роль воды оставалась неизвестной до тех пор, пока на нее не указал в 1808 Н.Соссюр. В своих очень точных экспериментах он измерял прирост сухого веса растения, растущего в горшке с землей, а также определял объем поглощенного диоксида углерода и выделенного кислорода. Соссюр подтвердил, что весь углерод, включенный растением в органические вещества, происходит из диоксида углерода. В то же время он обнаружил, что прирост сухого вещества растения был больше, чем разность между весом поглощенного диоксида углерода и весом выделенного кислорода. Поскольку вес почвы в горшке существенно не изменялся, единственным возможным источником увеличения веса следовало считать воду. Так было показано, что одним из реагентов в фотосинтезе является вода. Значение фотосинтеза как одного из процессов превращения энергии не могло быть оценено до тех пор, пока не возникло само представление о химической энергии. В 1845 Р.Майер пришел к выводу, что при фотосинтезе световая энергия переходит в химическую потенциальную энергию, запасаемую в его продуктах.


Роль фотосинтеза. Суммарный итог химических реакций фотосинтеза может быть описан для каждого из его продуктов отдельным химическим уравнением. Для простого сахара глюкозы уравнение имеет следующий вид:
Уравнение показывает, что в зеленом растении за счет энергии света из шести молекул воды и шести молекул диоксида углерода образуется одна молекула глюкозы и шесть молекул кислорода. Глюкоза - это лишь один из многих углеводов, синтезируемых в растениях. Ниже приведено общее уравнение для образования углевода с n атомами углерода в молекуле:
Уравнения, описывающие образование других органических соединений, имеют не столь простой вид. Для синтеза аминокислоты требуются дополнительные неорганические соединения, как например при образовании цистеина:
Роль света как реагента в процессе фотосинтеза легче доказать, если обратиться к другой химической реакции, а именно к горению. Глюкоза - одна из субъединиц целлюлозы, главного компонента древесины. Горение глюкозы описывается следующим уравнением:
Это уравнение представляет собой обращение уравнения фотосинтеза глюкозы, если не считать того, что вместо световой энергии выделяется главным образом тепло. По закону сохранения энергии, если при горении энергия выделяется, то при обратной реакции, т.е. при фотосинтезе, она должна поглощаться. Биологический аналог горения - дыхание, поэтому дыхание описывается тем же уравнением, что и небиологическое горение. Для всех живых клеток, за исключением клеток зеленых растений на свету, источником энергии служат биохимические реакции. Дыхание - главный биохимический процесс, высвобождающий энергию, запасенную в ходе фотосинтеза, хотя между этими двумя процессами могут лежать длинные пищевые цепи. Постоянный приток энергии необходим для любого проявления жизнедеятельности, и световая энергия, которую фотосинтез преобразует в химическую потенциальную энергию органических веществ и использует на выделение свободного кислорода, - это единственно важный первичный источник энергии для всего живого. Живые клетки затем окисляют ("сжигают") эти органические вещества с помощью кислорода, и часть энергии, высвободившейся при соединении кислорода с углеродом, водородом, азотом и серой, запасают для использования в различных процессах жизнедеятельности, таких, как движение или рост. Соединяясь с перечисленными элементами, кислород образует их оксиды - диоксид углерода, воду, нитрат и сульфат. Тем самым цикл завершается. Почему свободный кислород, единственным источником которого на Земле служит фотосинтез, так необходим для всего живого? Причина заключается в его высокой реакционной способности. В электронном облаке нейтрального атома кислорода на два электрона меньше, чем требуется для наиболее стабильной электронной конфигурации. Поэтому у атомов кислорода сильно выражена тенденция к приобретению двух дополнительных электронов, что достигается путем объединения (образования двух связей) с другими атомами. Атом кислорода может образовать две связи с двумя разными атомами или образовать двойную связь с одним каким-нибудь атомом. В каждой из таких связей один электрон поставляет атом кислорода, а второй электрон поставляется другим атомом, участвующим в образовании связи. В молекуле воды (Н2О), например, каждый из двух атомов водорода поставляет для образования связи с кислородом свой единственный электрон, удовлетворяя тем самым свойственное кислороду стремление к приобретению двух дополнительных электронов. В молекуле СО2 каждый из двух атомов кислорода образует двойную связь с одним и тем же атомом углерода, имеющим четыре связующих электрона. Таким образом, и в Н2О и в СО2 у атома кислорода столько электронов, сколько необходимо для стабильной конфигурации. Если, однако, два атома кислорода соединяются друг с другом, то электронные орбитали этих атомов допускают возникновение только одной связи. Потребность в электронах оказывается, таким образом, удовлетворена только наполовину. Поэтому молекула О2 по сравнению с молекулами СО2 и Н2О менее стабильна и более реакционноспособна. Органические продукты фотосинтеза, например углеводы, (СН2О)n, вполне стабильны, поскольку в них каждый из атомов углерода, водорода и кислорода получает столько электронов, сколько необходимо для образования наиболее стабильной конфигурации. Процесс фотосинтеза, в результате которого образуются углеводы, превращает, следовательно, два очень стабильных вещества, СО2 и Н2О, в одно вполне стабильное, (СН2О)n, и одно менее стабильное, О2. Накопление в результате фотосинтеза огромных количеств О2 в атмосфере и его высокая реакционная способность определяют его роль универсального окислителя. Когда какой-нибудь элемент отдает электроны или атомы водорода, мы говорим, что этот элемент окисляется. Присоединение электронов или образование связей с водородом, как у атомов углерода при фотосинтезе, называют восстановлением. Используя эти понятия, фотосинтез можно определить как окисление воды, сопряженное с восстановлением диоксида углерода или других неорганических оксидов.
Механизм фотосинтеза.
Световая и темновая стадии. В настоящее время установлено, что фотосинтез протекает в две стадии: световую и темновую. Световая стадия - это процесс использования света для расщепления воды; при этом выделяется кислород и образуются богатые энергией соединения. Темновая стадия включает группу реакций, в которых используются высокоэнергетические продукты световой стадии для восстановления СО2 до простого сахара, т.е. для ассимиляции углерода. Поэтому темновую стадию называют также стадией синтеза. Термин "темновая стадия" означает лишь то, что свет в ней непосредственно не участвует. Современные представления о механизме фотосинтеза сформировались на основе исследований, проведенных в 1930-1950-х годах. До этого на протяжении многих лет ученых вводила в заблуждение на первый взгляд простая, однако неверная гипотеза, согласно которой О2 образуется из СО2, а освободившийся углерод реагирует с Н2О, в результате чего и образуются углеводы. В 1930-х годах, когда выяснилось, что у некоторых серных бактерий кислород при фотосинтезе не выделяется, биохимик К. ван Ниль предположил, что кислород, выделяющийся в процессе фотосинтеза у зеленых растений, происходит из воды. У серных бактерий реакция протекает следующим образом:
Вместо О2 эти организмы образуют серу. Ван Ниль пришел к заключению, что все виды фотосинтеза можно описать уравнением
где Х - кислород в фотосинтезе, идущем с выделением О2, и сера в фотосинтезе серных бактерий. Ван Ниль также предположил, что этот процесс включает две стадии: световую и стадию синтеза. Эту гипотезу подкрепило открытие физиолога Р.Хилла. Он обнаружил, что разрушенные или частично инактивированные клетки способны на свету осуществлять реакцию, в которой кислород выделяется, но СО2 не восстанавливается (ее назвали реакцией Хилла). Чтобы эта реакция могла идти, требовалось добавить какой-нибудь окислитель, способный присоединять электроны или водородные атомы, отдаваемые кислородом воды. Один из реагентов Хилла - это хинон, который, присоединив два атома водорода, превращается в дигидрохинон. Другие реагенты Хилла содержали трехвалентное железо (ион Fe3+), которое, присоединив один электрон от кислорода воды, превращалось в двухвалентное (Fe2+). Так было показано, что переход водородных атомов от кислорода воды на углерод может совершаться в форме независимого движения электронов и ионов водорода. В настоящее время установлено, что для запасания энергии важен именно переход электронов от одного атома к другому, тогда как ионы водорода могут переходить в водный раствор, а при необходимости вновь из него извлекаться. Реакция Хилла, в которой световая энергия используется для того, чтобы вызвать перенос электронов от кислорода на окислитель (акцептор электронов), была первой демонстрацией перехода световой энергии в химическую и моделью световой стадии фотосинтеза. Гипотеза, согласно которой кислород во время фотосинтеза непрерывно поступает от воды, нашла дальнейшее подтверждение в опытах с применением воды, меченной тяжелым изотопом кислорода (18О). Поскольку изотопы кислорода (обычный 16О и тяжелый 18О) по своим химическим свойствам одинаковы, растения используют Н218О точно так же, как Н216О. Оказалось, что в выделенном кислороде присутствует 18О. В другом опыте растения вели фотосинтез с Н216О и С18О2. При этом выделяемый в начале эксперимента кислород не содержал 18О. В 1950-х годах физиолог растений Д.Арнон и другие исследователи доказали, что фотосинтез включает световую и темновую стадии. Из растительных клеток были получены препараты, способные осуществлять всю световую стадию. Используя их, удалось установить, что на свету происходит перенос электронов от воды к фотосинтетическому окислителю, который в результате этого становится донором электронов для восстановления диоксида углерода на следующей стадии фотосинтеза. Переносчиком электронов служит никотинамидадениндинуклеотидфосфат. Его окисленную форму обозначают НАДФ+, а восстановленную (образующуюся после присоединения двух электронов и иона водорода) - НАДФЧН. В НАДФ+ атом азота пятивалентный (четыре связи и один положительный заряд), а в НАДФЧН - трехвалентный (три связи). НАДФ+ принадлежит к т.н. коферментам. Коферменты совместно с ферментами осуществляют многие химические реакции в живых системах, но в отличие от ферментов в ходе реакции изменяются. Большая часть преобразованной световой энергии, запасаемой в световой стадии фотосинтеза, запасается при переносе электронов от воды к НАДФ+. Образовавшийся НАДФЧН удерживает электроны не столь прочно, как кислород воды, и может отдавать их в процессах синтеза органических соединений, расходуя накопленную энергию на полезную химическую работу. Значительное количество энергии запасается еще и другим способом, а именно в форме АТФ (аденозинтрифосфата). Он образуется в результате отнятия воды от неорганического иона фосфата (HPO42-) и органического фосфата, аденозиндифосфата (АДФ), согласно следующему уравнению:

АТФ - богатое энергией соединение, и для его образования необходимо поступление энергии от какого-то источника. В обратной реакции, т.е. при расщеплении АТФ на АДФ и фосфат, энергия высвобождается. Во многих случаях АТФ отдает свою энергию другим химическим соединениям в реакции, в которой водород замещается на фосфат. В представленной ниже реакции сахар (ROH) фосфорилируется, превращаясь в сахарофосфат:

В сахарофосфате заключено больше энергии, чем в нефосфорилированном сахаре, поэтому его реакционная способность выше. АТФ и НАДФЧН, образующиеся (наряду с О2) в световой стадии фотосинтеза, используются затем на стадии синтеза углеводов и других органических соединений из диоксида углерода.
Устройство фотосинтетического аппарата.
Световая энергия поглощается пигментами (так называют вещества, поглощающие видимый свет). У всех растений, осуществляющих фотосинтез, имеются различные формы зеленого пигмента хлорофилла, и, вероятно, во всех содержатся каротиноиды, окрашенные обычно в желтые тона. В высших растениях содержатся хлорофилл а (С55Н72О5N4Mg) и хлорофилл b (C55H70O6N4Mg), а также четыре основных каротиноида: b-каротин (С40Н56), лютеин (С40Н55О2), виолаксантин и неоксантин. Такое разнообразие пигментов обеспечивает широкий спектр поглощения видимого света, поскольку каждый из них "настроен" на свою область спектра. У некоторых водорослей набор пигментов приблизительно тот же, однако у многих из них имеются пигменты, несколько отличающиеся от перечисленных по своей химической природе. Все эти пигменты, как и весь фотосинтетический аппарат зеленой клетки, заключены в особые органеллы, окруженные мембраной, т.н. хлоропласты. Зеленая окраска растительных клеток зависит только от хлоропластов; остальные элементы клеток зеленых пигментов не содержат. Размеры и форма хлоропластов довольно сильно варьируют. Типичный хлоропласт напоминает по форме слегка изогнутый огурец размерами ок. 1 мкм в поперечнике и длиной ок. 4 мкм. В крупных клетках зеленых растений, таких, как клетки листа у большинства наземных видов, содержится много хлоропластов, а у мелких одноклеточных водорослей, например у Chlorella pyrenoidosa, имеется только один хлоропласт, занимающий большую часть клетки.
Познакомиться с очень сложным строением хлоропластов позволяет электронный микроскоп. Он дает возможность выявить гораздо более мелкие структуры, нежели те, какие видны в обычном световом микроскопе. В световом микроскопе нельзя различить частицы мельче 0,5 мкм. Разрешающая способность электронных микроскопов уже к 1961 позволяла наблюдать и в тысячу раз более мелкие частицы (порядка 0,5 нм). С помощью электронного микроскопа в хлоропластах выявлены очень тонкие мембранные структуры, т.н. тилакоиды. Это плоские мешочки, сомкнутые по краям и собранные в стопки, называемые гранами; на снимках граны похожи на стопки очень тонких блинов. Внутри мешочков имеется пространство - полость тилакоидов, а сами тилакоиды, собранные в граны, погружены в гелеобразную массу растворимых белков, заполняющую внутреннее пространство хлоропласта и называемую стромой. В строме содержатся также более мелкие и тонкие тилакоиды, которые соединяют друг с другом отдельные граны. Все тилакоидные мембраны состоят примерно из равного количества белков и липидов. Независимо от того, собраны они в граны или нет, именно в них сосредоточены пигменты и протекает световая стадия. Темновая стадия протекает, как принято считать, в строме.
Фотосистемы.
Хлорофилл и каротиноиды, погруженные в тилакоидные мембраны хлоропластов, собраны в функциональные единицы - фотосистемы, каждая из которых содержит примерно 250 молекул пигментов. Устройство фотосистемы таково, что из всех этих молекул, способных поглощать свет, только одна особым образом расположенная молекула хлорофилла а может использовать его энергию в фотохимических реакциях - она является реакционным центром фотосистемы. Остальные молекулы пигментов, поглощая свет, передают его энергию на реакционный центр; эти светособирающие молекулы называют антенными. Существует два типа фотосистем. В фотосистеме I специфическая молекула хлорофилла а, составляющая реакционный центр, имеет оптимум поглощения при длине световой волны 700 нм (обозначается P700; P - пигмент), а в фотосистеме II - при 680 нм (P680). Обычно обе фотосистемы работают синхронно и (на свету) непрерывно, хотя фотосистема I может работать и отдельно.
Превращения световой энергии.
Рассмотрение этого вопроса следует начать с фотосистемы II, где энергия света утилизируется реакционным центром P680. Когда в эту фотосистему поступает свет, его энергия возбуждает молекулу P680, и пара возбужденных, энергизованных электронов, принадлежащих этой молекуле, отрывается и переносится на молекулу акцептора (вероятно, хинона), обозначаемого буквой Q. Ситуацию можно представить себе таким образом, что электроны как бы подскакивают от полученного светового "толчка" и акцептор ловит их в каком-то верхнем положении. Если бы не акцептор, электроны вернулись бы в исходное положение (на реакционный центр), а высвобождающаяся при движении вниз энергия переходила бы в световую, т.е. тратилась бы на флуоресценцию. С этой точки зрения, акцептор электронов можно рассматривать как гаситель флуоресценции (отсюда его обозначение Q, от англ. quench - гасить).
Молекула P680, потеряв два электрона, окислилась, и для того, чтобы процесс на этом не прекратился, она должна восстановиться, т.е. получить два электрона из какого-либо источника. Таким источником служит вода: она расщепляется на 2Н+ и 1/2O2, отдавая два электрона на окисленный P680. Это светозависимое расщепление воды называется фотолизом. Ферменты, осуществляющие фотолиз, находятся на внутренней стороне мембраны тилакоидов, вследствие чего все ионы водорода накапливаются в полости тилакоидов. Важнейшим кофактором ферментов фотолиза служат атомы марганца. Переход двух электронов от реакционного центра фотосистемы на акцептор - это подъем "в гору", т.е. на более высокий энергетический уровень, и этот подъем обеспечивает энергия света. Далее в фотосистеме II пара электронов начинает поэтапный "спуск" от акцептора Q к фотосистеме I. Спуск происходит по электрон-транспортной цепи, очень сходной по организации с аналогичной цепью в митохондриях (см. также МЕТАБОЛИЗМ). В ее состав входят цитохромы, белки, содержащие железо и серу, медь-содержащий белок и другие компоненты. Постепенный спуск электронов от более энергизованного состояния к менее энергизованному сопряжен с синтезом АТФ из АДФ и неорганического фосфата. В результате энергия света не утрачивается, а запасается в фосфатных связях АТФ, которые могут быть использованы в процессах метаболизма. Образование АТФ в ходе фотосинтеза называют фотофосфорилированием. Одновременно с описанным процессом идет поглощение света в фотосистеме I. Здесь его энергия тоже используется на отрыв двух электронов от реакционного центра (P700) и передачу их на акцептор - железосодержащий белок. От этого акцептора через промежуточный переносчик (тоже белок, содержащий железо) оба электрона идут на НАДФ+, который в результате становится способным присоединить ионы водорода (образовавшиеся при фотолизе воды и сохранившиеся в тилакоидах) - и превращается в НАДФЧН. Что касается окислившегося в начале процесса реакционного центра P700, то он принимает два ("спустившихся") электрона из фотосистемы II, что возвращает его в исходное состояние. Суммарную реакцию световой стадии, протекающей при фотоактивации фотосистем I и II, можно представить следующим образом:
Общий энергетический выход потока электронов при этом составляет 1 молекулу АТФ и 1 молекулу НАДФН на 2 электрона. Путем сравнения энергии этих соединений с энергией света, обеспечивающего их синтез, было вычислено, что в процессе фотосинтеза запасается примерно 1/3 энергии поглощенного света. У некоторых фотосинтезирующих бактерий фотосистема I работает независимо. При этом поток электронов движется циклически от реакционного центра на акцептор и - по обходному пути - обратно на реакционный центр. В этом случае не происходит фотолиза воды и выделения кислорода, не образуется НАДФЧН, но АТФ синтезируется. Такой механизм световой реакции может иметь место и у высших растений в условиях, когда в клетках возникает избыток НАДФЧН.
Темновые реакции (стадия синтеза).
Синтез органических соединений путем восстановления СО2 (а также нитрата и сульфата) тоже происходит в хлоропластах. АТФ и НАДФЧН, поставляемые световой реакцией, протекающей в тилакоидных мембранах, служат для реакций синтеза источником энергии и электронов. Восстановление СО2 есть результат переноса электронов на СО2. В ходе этого переноса некоторые из связей С-О заменяются на связи С-Н, С-С и О-Н. Процесс состоит из ряда этапов, часть которых (15 или более) образует цикл. Этот цикл был открыт в 1953 химиком М.Калвином и его сотрудниками. Использовав в своих опытах вместо обычного (стабильного) изотопа углерода его радиоактивный изотоп, эти исследователи смогли проследить путь углерода в изучаемых реакциях. В 1961 Калвин был удостоен за эту работу Нобелевской премии по химии. В цикле Калвина участвуют соединения с числом атомов углерода в молекулах от трех до семи. Все компоненты цикла, за исключением одного, представляют собой сахарофосфаты, т.е. сахара, у которых одна или две ОН-группы заменены на фосфатную группу (-ОРО3Н-). Исключение составляет 3-фосфоглицериновая кислота (ФГК; 3-фосфоглицерат), представляющая собой фосфат сахарной кислоты. Она сходна с фосфорилированным трехуглеродным сахаром (глицерофосфатом), но отличается от него тем, что имеет карбоксильную группу O=C-O-, т.е. один из ее углеродных атомов соединен с атомами кислорода тремя связями. Начать описание цикла удобно с рибулозомонофосфата, содержащего пять атомов углерода (C5). Образующийся в световой стадии АТФ реагирует с рибулозомонофосфатом, превращая его в рибулозодифосфат. Вторая фосфатная группа придает рибулозодифосфату дополнительную энергию, поскольку несет в себе часть энергии, запасенной в молекуле АТФ. Поэтому тенденция реагировать с другими соединениями и образовывать новые связи выражена у рибулозодифосфата сильнее. Именно этот C5-сахар присоединяет CO2 с образованием шестиуглеродного соединения. Последнее очень неустойчиво и под действием воды распадается на два фрагмента - две молекулы ФГК. Если иметь в виду только изменение числа атомов углерода в молекулах сахаров, то этот основной этап цикла, в котором происходит фиксация (ассимиляция) CO2, можно представить следующим образом:

Фермент, катализирующий фиксацию CO2 (специфическая карбоксилаза), присутствует в хлоропластах в очень больших количествах (свыше 16% от общего содержания в них белка); учитывая огромную массу зеленых растений, он, вероятно, является самым распространенным белком в биосфере. Следующий этап состоит в том, что две молекулы ФГК, образовавшиеся в реакции карбоксилирования, восстанавливаются каждая за счет одной молекулы НАДФЧН до трехуглеродного сахарофосфата (триозофосфата). Это восстановление происходит в результате переноса двух электронов на углерод карбоксильной группы ФГК. Однако и в данном случае необходим АТФ, чтобы снабдить молекулу дополнительной химической энергией и повысить ее реакционную способность. Задачу эту выполняет ферментная система, которая переносит концевую фосфатную группу АТФ на один из атомов кислорода карбоксильной группы (образуется группа), т.е. ФГК превращается в дифосфоглицериновую кислоту. Как только НАДФЧН передает углероду карбоксильной группы этого соединения один атом водорода плюс электрон (что равноценно двум электронам плюс ион водорода, Н+), одинарная связь С-О разрывается и связанный с фосфором кислород переходит в неорганический фосфат, HPO42-, а карбоксильная группа O=C-O- превращается в альдегидную O=C-H. Последняя характерна для определенного класса сахаров. В итоге ФГК при участии АТФ и НАДФЧН восстанавливается до сахарофосфата (триозофосфата). Весь описанный выше процесс может быть представлен следующими уравнениями: 1) Рибулозомонофосфат + АТФ -> Рибулозодифосфат + АДФ 2) Рибулозодифосфат + СО2 -> Нестойкое С6-соединение 3) Нестойкое С6-соединение + Н2О -> 2 ФГК 4) ФГК + АТФ + НАДФЧН -> АДФ + H2PO42- + Триозофосфат (С3). Конечным результатом реакций 1-4 оказывается образование из рибулозомонофосфата и СО2 двух молекул триозофосфата (С3) с затратой двух молекул НАДФЧН и трех молекул АТФ. Именно в этой серии реакций представлен весь вклад световой стадии - в форме АТФ и НАДФЧН - в цикл восстановления углерода. Разумеется, световая стадия должна дополнительно поставлять эти кофакторы для восстановления нитрата и сульфата и для превращения ФГК и триозофосфата, образуемых в цикле, в другие органические вещества - углеводы, белки и жиры. Значение последующих этапов цикла сводится к тому, что они приводят к регенерации пятиуглеродного соединения, рибулозомонофосфата, необходимого для возобновления цикла. Эту часть цикла можно записать в следующем виде:

что дает в сумме 5С3 -> 3С5. Три молекулы рибулозомонофосфата, образовавшиеся из пяти молекул триозофосфата, превращаются - после присоединения CO2 (карбоксилирования) и восстановления - в шесть молекул триозофосфата. Таким образом, в результате одного оборота цикла одна молекула диоксида углерода включается в состав трехуглеродного органического соединения; три оборота цикла суммарно дают новую молекулу последнего, а для синтеза молекулы шестиуглеродного сахара (глюкозы или фруктозы) необходимы две трехуглеродные молекулы и соответственно 6 оборотов цикла. Прирост органического вещества цикл отдает реакциям, в которых образуются различные сахара, жирные кислоты и аминокислоты, т.е. "строительные блоки" крахмала, жиров и белков. Тот факт, что прямыми продуктами фотосинтеза являются не только углеводы, но также аминокислоты, а возможно и жирные кислоты, тоже был установлен с помощью изотопной метки - радиоактивного изотопа углерода. Хлоропласт - это не просто частица, приспособленная для синтеза крахмала и сахаров. Это весьма сложная, прекрасно организованная "фабрика", способная не только производить все материалы, из которых построена она сама, но и снабжать восстановленными соединениями углерода те части клетки и те органы растения, которые сами фотосинтеза не ведут.
ЛИТЕРАТУРА
Эдвардс Дж., Уокер Д. Фотосинтез C3- и C4-растений: механизмы и регуляция. М., 1986 Рейвн П., Эверт Р., Айкхорн С. Современная ботаника, т. 1. М., 1990
Энциклопедия Кольера. - Открытое общество . 2000 .