Строение синапса: электрические и химические синапсы. Областной университет синапс как функциональный контакт нервной ткани


5. Химические синапсы по природе медиатора делят на холин-эргические (медиатор - ацетилхолин), адренергические (норадре-налин), дофаминэргические (дофамин), ГАМК-эргические (у-аминомасляная кислота) и т. д. В ЦНС в основном химические синапсы, однако имеются и электрические возбуждающие си-напсы и электрохимические синапсы.
Б. Структурные элементы химического синапса - преси-наптическая и постсинаптическая мембраны и синаптическая щель (рис. 2.5).
В пресинаптическом окончании находятся синаптические пузырьки (везикулы) диаметром около 40 нм, которые образуются в теле нейрона и с помощью микротрубочек и микрофиламентов доставляются в пресин^птическое окончание, где заполняются ме-диатором и АТФ. Медиатор образуется в самом нервном оконча-нии. В пресинаптическом окончании содержатся несколько тысяч везикул, в каждой из которых от 1 до 10 тысяч молекул химическо-го вещества, участвующего в передаче влияния через синапс и в связи с этим названного медиатором (посредником). Митохондрии пресинаптического окончания обеспечивают энергией процесс си-наптической передачи. Пресинаптической мембраной называют часть мембраны пресинаптического окончания, ограничивающую синаптическую щель.
Синаптическая щель имеет различную ширину (20-50 нм), содержит межклеточную жидкость и мукополисахаридное плотное
вещество в виде полосок, мостиков, которое обеспечивает связь между пре- и постсинаптической мембранами и может содержать ферменты.
Постсинаптическая мембрана - это утолщенная часть кле-точной мембраны иннервируемой клетки, содержащая белковые рецепторы, имеющие ионные каналы и способные связывать моле-кулы медиатора. Постсинаптическую мембрану нервно-мышечно-го синапса называют также концевой пластинкой.
В. Механизм передачи возбуждения в электрическом си-напсе подобен таковому в нервном волокне: ПД, возникающий на пресинаптической мембране, непосредственно электрически раз-дражает постсинаптическую мембрану и обеспечивает возбужде-ние ее. Электрические синапсы, как выяснилось, оказывают опре-деленное влияние на метаболизм контактирующих клеток. Имеются данные о наличии в ЦНС и тормозных электрических си-напсов, однако они изучены недостаточно.
Г. Передача сигнала в химических синапсах. Потенциал действия (ПД), поступивший в пресинаптическое окончание хими-ческого синапса, вызывает деполяризацию его мембраны, открыва-ющую потенциалзависимые Са-каналы. Ионы Са 2+ входят внутрь нервного окончания согласно электрохимическому градиенту » обеспечивают выделение медиатора в синаптическую щель посред-ством экзоцитоза. Молекулы медиатора, поступившие в синапти-ческую щель, диффундируют к постсинаптической мембране и всту-пают во взаимодействие с ее рецепторами. Действие молекул медиатора ведет к открытию ионных каналов и перемещению ионов Ыа + и К + согласно электрохимическому градиенту с преобладани-ем тока ионов Ыа + в клетку, что ведет к ее деполяризации. Эта де-поляризация называется возбуждающим постсинаптическим потен-циалом (ВПСП), который в нервномышечном синапсе называют потенциалом концевой пластинки (ПКП) (рис. 2.6).
Прекращение действия медиатора, выделившегося в синапти-ческую щель, осуществляется с помощью его разрушения фермен-тами, локализующимися в синаптической щели и на постсинапти-ческой мембране, путем диффузии медиатора в окружающую среду, а также с помощью обратного захвата нервным окончанием.
Д. Характеристика проведения возбуждения в химиче-ских синапсах.
1 . Одностороннее проведение возбуждения - от пресинапти-ческого окончания в сторону постсинаптической мембраны. Это связано с тем, что медиатор выделяется из пресинаптического окон-чания, а взаимодействующие с ним рецепторы локализуются толь-ко на постсинаптической мембране.

Замедленное распространение возбуждения в синапсах по сравнению с нервным волокном объясняется тем, что необходимо время на выделение медиатора из пресинаптического окончания, распространение медиатора в синаптической щели, действие ме-диатора на постсинаптическую мембрану. Суммарная задержка передачи возбуждения в нейроне достигает величины порядка 2 мс, в нервно-мышечном синапсе 0,5-1,0 мс.
Низкая лабильность химических синапсов. В нервно-мы-шечном синапсе равна 100-150 передаваемым импульсам в секун-ду, что в 5-6 раз ниже лабильности нервного волокна. В синапсах ЦНС весьма вариабельна - может быть больше или меньше. При-чина низкой лабильности синапса - синаптическая задержка.
4. Синаптическая депрессия (утомляемость синапса) -
ослабление реакции клетки на афферентные импульсы, выражаю-
щееся в снижении постсинаптических потенциалов во время дли-
тельного раздражения или после него. Оно объясняется расходо-
ванием медиатора, накоплением метаболитов, закислением среды
при длительном проведении возбуждения по одним и тем же ней-
ронным цепям.
Е. Электрические синапсы имеют щель на порядок меньше, чем у химических синапсов, проводят сигнал в обе стороны без си-наптической задержки, передача не блокируется при удалении Са 2+ , они мало чувствительны к фармакологическим препаратам и ядам, практически неутомляемы, как и нервное волокно. Очень низкое удельное сопротивление сближенных пре- и постсинаптических мембран обеспечивает хорошую электрическую проводимость.
2.2. ХАРАКТЕРИСТИКА ГОРМОНАЛЬНОЙ РЕГУЛЯЦИИ
У рефлекторной реакции может быть гормональное зве-но, что характерно для регуляции функций внутренних органов -вегетативных функций, в отличие от соматических функций, реф-лекторная регуляция которых осуществляется только нервным пу-тем (деятельность опорно-двигательного аппарата). Если включа-ется гормональное звено, то это осуществляется за счет дополнительной выработки биологически активных веществ. Напри-мер, при действии на экстерорецепторы сильных раздражителей (холод, жара, болевой раздражитель) возникает мощный поток аф-ферентных импульсов, поступающих в ЦНС, при этом в кровь выб-расывается дополнительное количество адреналина и гормонов коры надпочечников, играющих адаптивную (защитную) роль.
Гормоны (греч. погтаб - возбуждаю) - биологически актив-ные вещества, вырабатываемые эндокринными железами или спе-циализированными клетками, находящимися в различных органах (например, в поджелудочной железе, в желудочно-кишечном трак-те). Гормоны вырабатываются также нервными клетками - ней-рогормоны, например, гормоны гипоталамуса (либерины и стати-ны), регулирующие функцию гипофиза. Биологически активные вещества вырабатываются также неспециализированными клет-ками - тканевые гормоны (паракринные гормоны, гормоны мест-ного действия, паракринные факторы - парагормоны). Действие гормонов или парагормонов непосредственно на соседние клет-ки, минуя кровь, называют паракринным действием. По месту действия на органы-мишени или на другие эндокринные железы гормоны делят на две группы: 1) эффекторные гормоны, дей-ствующие на клетки-эффекторы (например, инсулин, регулирую-щий обмен веществ в организме, увеличивает синтез гликогена в клетках печени, увеличивает транспорт глюкозы и других веществ через клеточную мембрану, повышает интенсивность синтеза бел-ка); 2) тройные гормоны (тропины), действующие на другие эндокринные железы и регулирующие их функции (например, ад-
ренокортикотропный гормон гипофиза - кортикотропин (АКТГ) -регулирует выработку гормонов корой надпочечников).
Виды влияний гормонов. Гормоны оказывают два вида влия-ний на органы, ткани и системы организма: функциональное (игра-ют весьма важную роль в регуляции функций организма) и морфоге-нетическое (обеспечивают морфогенез - рост, физическое, половое и умственное развитие; например, при недостатке тироксина стра-дает развитие ЦНС, следовательно, и умственное развитие).
1. Функциональное влияние гормонов бывает трех видов.
Пусковое влияние - это способность гормона запускать дея-тельность эффектора. Например, адреналин запускает распад гли-когена в печени и выход глюкозы в кровь, вазопрессин (антидиуре-тический гормон - АДГ) включает реабсорбцию воды из собирательных трубок нефрона в интерстиций почки.
Модулирующее влияние гормона - изменение интенсивности протекания биохимических процессов в органах и тканях. Напри-мер, активация тироксином окислительных процессов, которые могут проходить и без него; стимуляция адреналином деятельнос-ти сердца, которая проходит и без адреналина. Модулирующим влиянием гормонов является также изменение чувствительности ткани к действию других гормонов. Например, фолликулин усили-вает действие прогестерона на слизистую оболочку матки, тирео-идные гормоны усиливают эффекты катехоламинов.
Пермиссивное влияние гормонов - способность одного гормо-на обеспечивать реализацию эффекта другого гормона. Например, инсулин необходим для проявления действия соматотропного гор-мона, фоллитропин необходим для реализации эффекта лютропина.
2. Морфогенетическое влияние гормонов
(на рост, физическое
и половое развитие) подробно изучается другими дисциплинами
(гистология, биохимия) и лишь частично - в курсе физиологии (см.
гл. 6). Оба вида влияний гормонов (морфогенетическое и функцио-
нальное) реализуются сломощью метаболических процессов, за-
пускаемых посредством клеточных ферментных систем.
2.3. РЕГУЛЯЦИЯ С ПОМОЩЬЮ МЕТАБОЛИТОВ
И ТКАНЕВЫХ ГОРМОНОВ.
МИОГЕННЫЙ МЕХАНИЗМ РЕГУЛЯЦИИ.
РЕГУЛИРУЮЩАЯ ФУНКЦИЯ ГЭБ
Метаболиты - продукты, образующиеся в организме в процессе обмена веществ как результат различных биохимических реакций. Это аминокислоты, нуклеотиды, коферменты, угольная кислота, мо-
лочная, пировиноградная, адениловая кислоты, ионный сдвиг, изме-нения рН. Регуляция с помощью метаболитов на ранних этапах фило-генеза была единственной. Метаболиты одной клетки непосредствен-но влияли на другую, соседнюю клетку или группу клеток, которые в свою очередь таким же способом действовали на следующие клет-ки (контактная регуляция). С появлением гемолимфы и сосуди-стой системы метаболиты стали передаваться и другим клеткам орга-низма с движущейся гемолимфой на большие расстояния, причем осуществляться это стало быстрее. Затем появилась нервная систе-ма как регулирующая система, а еще позже - эндокринные желе-зы. Метаболиты хотя и действуют в основном как местные регуля-торы, но могут влиять и на другие органы и ткани, на активность нервных центров. Например, накопление угольной кислоты в крови ведет к возбуждению дыхательного центра и усилению дыхания. Примером местной гуморальной регуляции может служить гипере-мия интенсивно работающей скелетной мышцы - накапливающие-ся метаболиты обеспечивают расширение кровеносных сосудов, что увеличивает доставку кислорода и питательных веществ к мышце. Подобные регуляторные влияния метаболитов происходят и в дру-гих активно работающих органах и тканях организма.
Тканевые гормоны: биогенные амины (гистамин, серотонигг), простагландины и кинины. Занимают промежуточное положение между гормонами и метаболитами как гуморальные факторы ре-гуляции. Эти вещества свое регулирующее влияние оказывают на клетки тканей посредством изменения их биофизических свойств (проницаемости мембран, их возбудимости), изменения интенсивности обменных процессов, чувствительности клеточных рецепторов, образования вторых посредников. В результате это-го изменяется чувствительность клеток к нервным и гумораль-ным влияниям. Поэтому тканевые гормоны называют модуля-торами регуляторных сигналов - они оказывают модулирующее влияние. Тканевые гормоны образуются неспециализированны-ми клетками, но действуют они посредством специализированных клеточных рецепторов, например, для гистамина обнаружено два вида рецепторов - Н (и Н 2 . Поскольку тканевые гормоны влияют на проницаемость клеточных мембран, они регулируют поступ-ление в клетку и выход из клетки различных веществ и ионов, определяющих мембранный потенциал, а значит и развитие по-тенциала действия.
Миогенный механизм регуляции. С развитием мышечной системы в процессе эволюции миогенный механизм регуляции фун-кций постепенно становится все более заметным. Организм чело-века примерно на 50% состоит из мышц. Это скелетная мускулату-
ра (40% массы тела), мышца сердца, гладкие мышцы кровеносных и лимфатических сосудов, стенки желудочно-кишечного тракта, желчного, мочевого пузырей и других внутренних органов.
Сущность миогенного механизма регуляции состоит в том, что предварительное умеренное растяжение скелетной или сердечной мышцы увеличивает силу их сокращений. Сократительная актив-ность гладкой мышцы также зависит от степени наполнения поло-го мышечного органа, а значит и его растяжения. При увеличении наполнения органа тонус гладкой мышцы сначала возрастает, а за-тем возвращается к исходному уровню (пластичность гладкой мыш-цы), что обеспечивает регуляцию тонуса сосудов и наполнение внут-ренних полых органов без существенного повышения давления в них (до определенной величины). Кроме того, большинство глад-ких мышц обладают автоматией, они постоянно находятся в неко-торой степени сокращения под влиянием импульсов, возникающих в них самих (например, мышцы кишечника, кровеносных сосудов). Импульсы, поступающие к ним по вегетативным нервам, оказыва-ют модулирующее влияние - увеличивают или уменьшают тонус гладких мышечных волокон.
Регулирующая функция ГЭБ заключается и в том, что он формирует особую внутреннюю среду мозга, обеспечивающую оп-тимальный режим деятельности нервных клеток. Считают, что ба-рьерную функцию при этом выполняет особая структура стенок капилляров мозга. Их эндотелий имеет очень мало пор, узкие ще-левые контакты между клетками почти не содержат окошек. Со-ставной частью барьера являются также глиальные клетки, обра-зующие своеобразные футляры вокруг капилляров, покрывающие около 90% их поверхности. Наибольший вклад в развитие пред-ставлений о гемато-энцефалическом барьере сделали Л. С. Штерн и ее сотрудники. Этот барьер пропускает воду, ионы, глюкозу, амино-кислоты, газы, задерживая многие физиологически активные веще-ства: адреналин, серотонин, дофамин, инсулин, тироксин. Однако в нем существуют «окна»,*через которые соответствующие клетки мозга - хеморецепторы - получают прямую информацию о наличии в крови гормонов и других, не проникающих через барьер веществ; клетки мозга выделяют и свои нейросекреты. Зоны мозга, не имею-щие собственного гемато-энцефалического барьера, - это гипофиз, эпифиз, некоторые отделы гипоталамуса и продолговатого мозга.
ГЭБ выполняет также защитную функцию - предотвра-щает попадание микробов, чужеродных или токсических веществ экзо- и эндогенной природы в межклеточные пространства мозга. ГЭБ не пропускает многие лекарственные вещества, что необходи-мо учитывать в медицинской практике.
2.4. СИСТЕМНЫЙ ПРИНЦИП РЕГУЛЯЦИИ
Поддержание показателей внутренней среды организма осуще-ствляется с помощью регуляции деятельности различных органов и физиологических систем, объединяемых в единую функциональ-ную систему - организм. Представление о функциональных систе-мах разработал П. К. Анохин (1898-1974). В последние годы тео-рия функциональных систем успешно развивается К. В. Судаковым.
А. Структура функциональной системы. Функциональная система - это динамическая совокупность различных органов и физиологических систем организма, формирующаяся для достиже-ния полезного приспособительного результата. Например, чтобы быстро пробежать дистанцию, необходимо максимально усилить деятельность сердечнососудистой, дыхательной, нервной систем и мышц. Функциональная система включает следующие элементы: 1) управляющее устройство - нервный центр, представляющий объединение ядер различных уровней ЦНС; 2) его выходные ка-налы (нервы и гормоны); 3) исполнительные органы - эффекто- ры, обеспечивающие в ходе физиологической деятельности поддер-жание регулируемого процесса (показателя) на некотором оптимальном уровне (полезный результат деятельности функцио-нальной системы); 4) рецепторы результата (сенсорные рецеп-торы) - датчики, воспринимающие информацию о параметрах от-клонения регулируемого процесса (показателя) от оптимального уровня; 5) канал обратной связи (входные каналы), информиру-ющий нервный центр с помощью импульсаций от рецепторов ре-зультата или с помощью непосредственного действия химических веществ на центр - информация о достаточности или недостаточ-ности эффекторных усилий по поддержанию регулируемого про-цесса (показателя) на оптимальном уровне (рис. 2.7).
Афферентные импульсы от рецепторов результата по каналам обратной связи поступают в нервный центр, регулирующий тот или иной показатель, центр обеспечивает изменение интенсивности работы соответствующего органа.
При изменении интенсивности работы эффектора изменяется интенсивность метаболизма, что также играет важную роль в регуляции деятельности органов той или иной функциональной системы (гуморальный процесс регуляции).
Б. Мультипараметрический принцип взаимодействия различных функциональных систем - принцип, определяющий обобщенную деятельность функциональных систем (К. В. Судаков). Относительная стабильность показателей внутренней среды орга-низма является результатом согласованной деятельности многих

функциональных систем. Выяснилось, что различные показатели внутренней среды организма оказываются взаимосвязанными. Например, избыточное поступление воды в организм сопровожда-ется увеличением объема циркулирующей крови, повышением артериального давления, снижением осмотического давления плаз-мы крови. В функциональной системе, поддерживающей оптималь-ный уровень газового состава крови, одновременно осуществляет-ся взаимодействие рН, Р С02 и Р 02 . Изменение одного из этих параметров немедленно приводит к изменению количественных ха-рактеристик других параметров. Для достижения любого приспо-собительного результата формируется соответствующая функцио-нальная система.
В. Системогенез. Согласно П. К. Анохину, системогенез -избирательное созревание и развитие функциональных систем в анте- и постнатальнрм онтогенезе. В настоящее время тер-мин «системогенез» применяется в более широком смысле, при этом под системогенезом понимают не только процессы онтогенетиче-ского созревания функциональных систем, но и формирование и преобразование функциональных систем в ходе жизнедеятельнос-ти организма.
Системообразующими факторами функциональной системы любого уровня являются полезный для жизнедеятельности организ-ма приспособительный результат, необходимый в данный момент, и формирующаяся при этом мотивация. Например, для соверше-ния прыжка в высоту с шестом ведущую роль играют мышцы верх-
них конечностей, при прыжке в длину - мышцы нижних конечно-стей.
Гетерохронность созревания функциональных систем. В ходе антенатального онтогенеза различные структуры организма зак-ладываются в разное время и созревают различными темпами. Так, нервный центр группируется и созревает обычно раньше, чем закла-дывается и созревает иннервируемый им субстрат. В онтогенезе созревают в первую очередь те функциональные системы, без которых невозможно дальнейшее развитие организма. Напри-мер, из трех функциональных систем, связанных с полостью рта, пос-ле рождения сформированной оказывается лишь функциональная система сосания, позже формируется функциональная система жевания, затем функциональная система речи.
Консолидация компонентов функциональной системы - объединение в функциональную систему отдельных фрагментов, развивающихся в различных частях организма. Консолидация фраг-ментов функциональной системы - критический пункт развития ее физиологической архитектуры. Ведущую роль в этом про-цессе играет ЦНС. Например, сердце, сосуды, дыхательный аппа-рат, кровь объединяются в функциональную систему поддержания постоянства газового состава внутренней среды на основе совер-шенствования связей между различными отделами ЦНС, а также на основе развития иннервационных связей между ЦНС и соответ-ствующими периферическими структурами.
Все функциональные системы различного уровня имеют одинаковую архитектонику (структуру).
2.5. ТИПЫ РЕГУЛЯЦИИ ФУНКЦИЙ ОРГАНИЗМА
1. Регуляция по отклонению - циклический механизм, при котором всякое отклонение от оптимального уровня регулируемого показателя мобилизует все аппараты функциональной системы к восстановлению его на прежнем уровне. Регуляция по отклонению предполагает наличие в составе системного комплекса канала отрицательной обратной связи, обеспечивающего разно-направленное влияние: усиление стимулирующих механизмов управления в случае ослабления показателей процесса или ослабление стимулирующих механизмов в случае чрезмерного усиления показателей процесса. Например, при повышении АД включаются регуляторные механизмы, обеспечивающие снижение АД, а при низком АД включаются противоположные реакции. В отличие от отрицательной обратной связи, положительная
обратная связь, встречающаяся в организме редко, оказывает только однонаправленное, усиливающее влияние на развитие процесса, находящегося под контролем управляющего комплекса. Поэтому положительная обратная связь делает систему неустой-чивой, неспособной обеспечить стабильность регулируемого процесса в пределах физиологического оптимума. Например, если бы артериальное давление регулировалось по принципу положи-тельной обратной связи, в случае снижения артериального давления действие регуляторных механизмов привело бы к еще большему его снижению, а в случае повышения - к еще большему его увеличению. Примером положительной обратной связи явля-ется усиление начавшейся секреции пищеварительных соков в желудке после приема пищи, что осуществляется с помощью продуктов гидролиза, всосавшихся в кровь.
2. Регуляция по опережению заключается в том, что регули-рующие механизмы включаются до реального изменения парамет-ра регулируемого процесса (показателя) на основе информации, поступающей в нервный центр функциональной системы и сигна-лизирующей о возможном изменении регулируемого процесса в будущем. Например, терморецепторы (детекторы температуры), находящиеся внутри тела, обеспечивают контроль за температурой внутренних областей тела. Терморецепторы кожи, в основном, иг-рают роль детекторов температуры окружающей среды. При зна-чительных отклонениях температуры окружающей среды создают-ся предпосылки возможного изменения температуры внутренней среды организма. Однако в норме этого не происходит, так как им-пульсация от терморецепторов кожи, непрерывно поступая в гипо-таламический терморегуляторный центр, позволяет ему произве-сти изменения работы эффекторов системы до момента реального изменения температуры внутренней среды орга-низма. Усиление вентиляции легких при физической нагрузке на-чинается раньше увеличения потребления кислорода и накопления угольной кислоты в крови человека. Это осуществляется благода-ря афферентной импульсации от проприорецепторов активно ра-ботающих мышц. Следовательно, импульсация проприорецепторов выступает как фактор, организующий перестройку работы функ-циональной системы, поддерживающей оптимальный для метабо-лизма уровень Р 02 , Р сс, 2 и рН внутренней среды с опережением.
Регуляция по опережению может реализоваться с помощью механизма условного рефлекса. Показано, что у кондукторов то-варных поездов в зимнее время резко нарастает производство теп-ла по мере удаления от станции отправления, где кондуктор нахо-дился в теплой комнате. На обратном пути, по мере приближения
физического
Что такое синапс? Синапс – это особая структура, предоставляющая передачу сигнала от волокон нервной клетки на прочую клетку или волокно от контактной клетки. Для чего необходимо наличие 2 нервных клеток. При этом синапс представлен в 3 функциональных участках (предсинаптический фрагмент, синаптическая щель и постсинаптический фрагмент) нервных клеток и располагается в области, где клетка контактирует с мышцами и железами человеческого организма.
 Система нейронных синапсов осуществляется по их локализации, типу деятельности и методу транзита имеющихся сигнальных данных. Относительно локализации синапсы различают: нейронейрональные, нервно-мышечные
. Нейронейрональные на аксосоматические, дендросоматические, аксодендритические, аксоаксональные.
Система нейронных синапсов осуществляется по их локализации, типу деятельности и методу транзита имеющихся сигнальных данных. Относительно локализации синапсы различают: нейронейрональные, нервно-мышечные
. Нейронейрональные на аксосоматические, дендросоматические, аксодендритические, аксоаксональные.
По типу деятельности на восприятие синапсы принято выделять: возбуждающие и не менее важные тормозящие. Относительно метода транзита информационного сигнала классифицируют их на:
- Электрический тип.
- Химический тип.
- Смешанный тип.
Этиология контактирования нейронов сводится к типу этого стыкования , которое может быть дистантным, контактным, а также пограничным. Соединение дистантного свойства выполняется посредством 2 нейронов, размещённых во многих частях организма.
Так, в тканях человеческого мозга генерируются нейрогормоны и вещества-нейропептиды, влияющие на присутствующие нейроны организма другого местоположения. Контактное соединение сводится к особым стыковкам плёнок-мембран типичных нейронов, составляющих синапсы химического направления, а также составляющих электрического свойства.
Смежная (пограничная) работа нейронов производится во время, в течение которого плёнки-мембраны нейронов перегорожены лишь синаптической щелью. Как правило, такое слияние наблюдается, если между 2 специальными плёнками-мембранами отсутствуют глиальные ткани . Данная смежность свойственна параллельным волокнам мозжечка, аксонам специального нерва обонятельного назначения и так далее.
Существует мнение, что смежный контакт провоцирует работу рядом расположенных нейронов в произведении общей функции. Это наблюдается по причине того, что метаболиты, плоды действия человеческого нейрона, проникая внутрь полости, расположенной, между клетками оказывают влияние на близлокализующиеся активные нейроны. Причём пограничное соединение часто может передавать данные электрического характера от 1 рабочего нейрона к 2 участнику процесса.
Синапсы электрического и химического направления
 Действие слияния плёнок-мембран принято считать электрическими синапсами
. В условиях, когда необходимая синаптическая щель прерывистая с промежутками перегородок монолитного соединения. Эти перегородки формируют чередующуюся конструкцию отделений синапса, при этом отделения обособлены фрагментами приближенных мембран, промежуток между которыми в синапсах обычного склада равен 0,15 - 0,20 нм у представителей млекопитающих существ. В месте соединения плёнок-мембран присутствуют пути, с помощью которых происходит обмен частью плодов.
Действие слияния плёнок-мембран принято считать электрическими синапсами
. В условиях, когда необходимая синаптическая щель прерывистая с промежутками перегородок монолитного соединения. Эти перегородки формируют чередующуюся конструкцию отделений синапса, при этом отделения обособлены фрагментами приближенных мембран, промежуток между которыми в синапсах обычного склада равен 0,15 - 0,20 нм у представителей млекопитающих существ. В месте соединения плёнок-мембран присутствуют пути, с помощью которых происходит обмен частью плодов.
Помимо отдельчатых типов синапсов существуют необходимые электрические типичные синапсы в виде единой синаптической щели, общий периметр которой простирается на 1000 мкм. Так, подобное синаптическое явление представлено в нейронах ресничного ганглия .
Электрические синапсы способны проводить качественное возбуждение в одностороннем порядке. Этот факт отмечается при фиксации электрического резерва синаптической составляющей. Например, в момент при касании афферентных канальцев синаптическая плёнка-мембрана деполяризуется, когда с касанием эфферентных частиц волокон напортив - гиперполяризуется. Считается, что синапсы действующих нейронов с общими обязанностями могут осуществлять требуемое возбуждение (между 2 пропускающими участками) в обе стороны.
Напротив, синапсы присутствующих нейронов с разным перечнем действий (моторные и сенсорные) проводят акт возбуждения односторонне . Основная работа синаптических составляющих обуславливается продуцированием безотлагательных реакций организма. Электрический синапс подлежит незначительной доли утомляемости, обладает значительным процентом устойчивости к внутренне-наружным факторам.
Химические синапсы обладают видом предсинаптического сегмента, функциональной синаптической щели с фрагментом постсинаптической составляющей. Предсинаптический фрагмент формируется увеличением размера аксона внутри собственного канальца или к его завершению. В этом фрагменте присутствуют гранулярные, а также агранулярные специальные мешочки, содержащие медиатор.
Предсинаптическое увеличение наблюдает локализацию активных митохондрий, генерирующую частицы вещества-гликогена, а также требуемую выработку медиатора и другое. В условиях частого соприкосновения с предсинаптическим полем резерв медиатора в имеющихся мешочках утрачивается.
Существует мнение, что малые гранулярные пузырьки имеют такое вещество, как норадреналин, а большие – катехоламины. Причём в агранулярных полостях (пузырьках) располагается ацетилхонин. Помимо этого, медиаторами усиленного возбуждения считаются вещества, образованные по типу вырабатываемой аспарагиновой или не менее значимой кислоты глутамина.
Действующие контакты синапса часто располагаются между:
- Дендритом и аксоном.
- Сомой и аксоном.
- Дендритами.
- Аксонами.
- Сомой клетки и дендритами.
Влияние выработанного медиатора относительно присутствующей постсинаптической плёнки-мембраны происходит из-за чрезмерного проникновения её частиц натрия. Генерация мощных изливаний частиц натрия из рабочей синаптической щели сквозь постсинаптическую плёнку-мембрану формирует её деполяризацию, образуя возбуждение постсинаптического резерва. Транзиту химического направления данных синапса свойственно синаптическое приостановление возбуждения по времени равному 0,5 мс с выработкой постсинаптического резерва, как реакция на предсинаптический поток.
Данная возможность в момент возбуждения представляется в деполяризации постсинаптической плёнки-мембраны, а в момент приостановления в её гиперполяризации. Из-за чего наблюдается приостановленный постсинаптический резерв . Как правило, во время сильного возбуждения повышается уровень проницаемости постсинаптической плёнки-мембраны.
Требуемое возбуждающее свойство фиксируется внутри нейронов, если в типичных синапсах работает норадреналин, вещество-дофамин, ацетил холин, важный серотонин, вещество Р и кислота глутамина.
Сдерживающий потенциал формируется во время влияния на синапсы из гамма-аминомаслянной кислоты и глицина.
Умственная работоспособность детей
 Работоспособность человека напрямую определяет его возраст, когда все значения увеличиваются одновременно с развитием и физическим ростом детей.
Работоспособность человека напрямую определяет его возраст, когда все значения увеличиваются одновременно с развитием и физическим ростом детей.
Точность и скорость умственных действий с возрастом осуществляется неравномерно в зависимости от прочих факторов, фиксирующих развитие и физический рост организма. Учащимся любого возраста, у которых присутствуют отклонения здоровья , характерна работоспособность низкого значения относительно окружающих крепких детей.
У здоровых первоклашек со сниженной готовностью организма к постоянному процессу обучения по некоторым показателям способность к действию является низкой, что усложняет борьбу с возникающими проблемами в процессе учёбы.
Скорость наступления ослабленности обуславливается исходным состоянием детской системы чувствительного нервного генеза, рабочим темпом и объёмом нагрузки. При этом дети склонны к переутомлению во время продолжительной неподвижности и когда выполняемые действия ребёнку неинтересны. После перерыва работоспособность становится прежней или становится выше прежней, причём лучше отдых делать не пассивным, но активным, переключившись на отличное от этого занятие.
 Первая часть учебного процесса у обычных детей начальных классов сопровождается отличной работоспособностью, но к окончанию 3 урока у них отмечается снижение концентрации внимания:
Первая часть учебного процесса у обычных детей начальных классов сопровождается отличной работоспособностью, но к окончанию 3 урока у них отмечается снижение концентрации внимания:
- Они глядят в окно.
- Невнимательно слушают слова учителя.
- Изменяют положения своего тела.
- Начинают разговаривать.
- Встают со своего места.
Специфически велики значения работоспособности у старшеклассников, обучающихся во 2 смену. Особенно важно обратить внимание на то, что достаточно коротко время для подготовки к занятиям до времени начала учебного действия в классе и не гарантирует полноценного избавления от пагубных изменений в центральной нервной системе. Умственная активность быстро истощается в первые часы уроков, что явно отмечается в отрицательном поведении.
Посему качественные сдвиги работоспособности наблюдаются у учеников младшего блока на уроках с 1 - 3, а блоках среднего-старшего звена на 4 - 5 занятии. В свою очередь, 6 урок проходит в условиях особенно сниженной способности к действию. При этом продолжительность занятия у 2 - 11 классников – 45 минут, что ослабляет состояние детей. Поэтому рекомендуется периодически сменять вид работы, а в середине урока провести активную паузу.
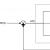
В большинстве синапсов нервной системы для передачи сигналов от пресинаптического нейрона на постсинаптический используются химические вещества - медиаторы или нейротрансмиттеры. Химическая передача сигналов осуществляется посредством химических синапсов (рис. 14), включающих мембраны пре- и постсинаптических клеток и разделяющую их синаптическую щель - область внеклеточного пространства шириной около 20 нм.
Рис.14. Химический синапс
В области синапса аксон обычно расширяется, образуя т.н. пресинаптическую бляшку или концевую пластинку. В пресинаптическом окончании содержатся синаптические везикулы - окруженные мембраной пузырьки диаметром порядка 50 нм, в каждом из которых заключено 10 4 – 5х10 4 молекул медиатора. Синаптическая щель заполнена мукополисахаридом, склеивающим между собой пре- и постсинаптические мембраны.
Установлена следующая последовательность событий при передаче через химический синапс. При достижении потенциалом действия пресинаптического окончания происходит деполяризация мембраны в зоне синапса, активируются кальциевые каналы плазматической мембраны и в окончание входят ионы Ca 2+ . Повышение внутриклеточного уровня кальция инициирует экзоцитоз везикул, наполненных медиатором. Содержимое везикул высвобождается во внеклеточное пространство, и часть молекул медиатора, диффундируя, связываются с рецепторными молекулами постсинаптической мембраны. Среди них есть рецепторы, которые могут непосредственно управлять ионными каналами. Связывание с такими рецепторами молекул медиатора является сигналом для активации ионных каналов. Таким образом, наряду с рассмотренными в предыдущем разделе потенциал-зависимыми ионными каналами существуют медиатор-зависимые каналы (иначе называются лиганд-активируемые каналы или ионотропные рецепторы). Они открываются и пропускают в клетку соответствующие ионы. Движение ионов по их электрохимическим градиентам, порождает натриевый деполяризующий (возбуждающий) или калиевый (хлорный) гиперполяризующий (тормозной) ток. Под воздействием деполяризующего тока развивается постсинаптический возбуждающий потенциал или потенциал концевой пластинки (ПКП). Если этот потенциал превосходит пороговый уровень, открываются потенциалзависимые натриевые каналы и возникает ПД. Скорость проведения импульса в синапсе меньше чем по волокну, т.е. наблюдается синаптическая задержка, напр., в нервно-мышечном синапсе лягушки – 0,5 мс. Описанная выше последовательность событий характерна для т.н. прямой синаптической передачи .
Кроме рецепторов непосредственно управляющих ионными каналами в химической передаче участвуют рецепторы, сопряженные с G-белками или метаботропные рецепторы .
G-белки, названные так по их способности связываться с гуаниновыми нуклеотидами, являются тримерами, состоящими из трех субъединиц: α, β и γ. Существует большое количество разновидностей каждой из субъединиц (20 α, 6 β, 12 γ). что создает основу для огромного количества их комбинаций. G-белки разделяются на четыре основных группы по структуре и мишеням их α-субъединиц: G s стимулирует аденилатциклазу; G i интибирует аденилатциклазу; G q связывается с фосфолипазой С; мишени С 12 пока не известны. Семейство G i включает G t (трансдуцин), который активирует фосфодиэстеразу цГМФ, а также две изоформы G 0 , которые связываются с ионными каналами. Вместе с тем, каждый из G-белков может взаимодействовать с несколькими эффекторами, а разные G-белки могут модулировать активность одних и тех же ионных каналов. В неактивированном состоянии гуанозиндифосфат (ГДФ) связан с α-субъединицей, и все три субъединицы объединены в тример. Взаимодействие с активированным рецептором позволяет гуанозинтрифосфату (ГТФ) заместить ГДФ на α-субъединице, что приводит к диссоциации α-- и βγ -субъединиц (в физиологических условиях β- и γ-субъединицы остаются связанными). Свободные α--и βγ -субъединицы связываются с белками-мишенями и модулируют их активность. Свободная α-субъединица обладает ГТФ-азной активностью, вызывая гидролиз ГТФ с образованием ГДФ. В результате α-- и βγ -субъединицы вновь связываются, что приводит к прекращению их активности.
В настоящее время идентифицировано > 1000 метаботропных рецепторов. В то время как рецепторы, связанные с каналами, вызывают электрические изменения в постсинаптической мембране всего за несколько миллисекунд или еще быстрее, рецепторам, не связанным с каналами, для достижения эффекта требуется несколько сотен миллисекунд и более. Это обусловлено тем обстоятельством, что между первоначальным сигналом и ответом должна пройти серия ферментативных реакций. Более того, часто сам сигнал "размыт" не только во времени, но и пространстве, поскольку установлено, что медиатор может высвобождаться не из нервных окончаний, а из варикозных утолщений (узелков), расположенных вдоль аксона. В этом случае отсутствуют морфологически выраженные синапсы, узелки не прилегают к каким-то специализированным рецептивным участкам постсинаптической клетки. Поэтому медиатор диффундирует в значительном объеме нервной ткани, воздействуя (как гормон) сразу на рецепторное поле у множества нервных клеток, расположенных в различных участках нервной системы и даже за ее пределами. Это т.н. непрямая синаптическая передача.
В ходе функционирования синапсы подвергаются функциональным и морфологическим перестройкам. Этот процесс назван синаптической пластичностью . Наиболее ярко такие изменения проявляются при высокочастотной активности, являющейся естественным условием функционирования синапсов in vivo. Например, частота импульсации вставочных нейронов в ЦНС достигает 1000 Гц. Пластичность может проявляться либо в увеличении (потенциации), либо уменьшении (депрессии) эффективности синаптической передачи. Выделяют кратковременные (длятся секунды и минуты) и долговременные (длятся часы, месяцы, годы) формы синаптической пластичности. Последние особенно интересны тем, что они имеют отношение к процессам научения и памяти. Например, долговременная потенциация - устойчивое усиление синаптической передачи в ответ на высокочастотное раздражение. Этот вид пластичности может продолжаться дни и месяцы. Долговременная потенциация наблюдается во всех отделах ЦНС, но наиболее полно изучена на глутаматергических синапсах в гиппокампе. Долговременная депрессия также возникает в ответ на высокочастотное раздражение и проявляется в виде длительного ослабления синаптической передачи. Этот вид пластичности имеет сходный механизм с долговременной потенциацией, но развивается при низкой внутриклеточной концентрации ионов Са2+, в то время как долговременная потенциация – при высокой.
На выделение медиаторов из пресинаптического окончания и химическую передачу нервного импульса в синапсе могут влиять медиаторы, выделяемые из третьего нейрона. Такие нейроны и медиаторы могут тормозить синаптическую передачу или, напротив, облегчать ее. В этих случаях говорят об гетеросинаптической модуляции - гетеросинаптическом торможении или облегчении в зависимости от конечного результата.
Таким образом, химическая передача более гибкая, чем электрическая, поскольку при этом без труда может осуществляться как возбуждающее, так и тормозное действие. Кроме того, при активации постсинаптических каналов химическими агентами может возникать достаточно сильный ток, способный деполяризовать крупные клетки.
Медиаторы - точки приложения и характер действия
Одна из самых сложных задач, стоящих перед нейрофизиологами, состоит в точной химической идентификации медиаторов, действующих в различных синапсах. К настоящему времени известно довольно много соединений, которые могут выполнять роль химических посредников при межклеточной передаче нервного импульса. Однако точно идентифицировано лишь ограниченное число таких медиаторов; некоторые из будут рассмотрены ниже. Для того чтобы медиаторная функция того или иного вещества в какой-либо ткани была неопровержимо доказана, должны удовлетворяться определенные критерии:
1. при прямом нанесении на постсинаптическую мембрану вещество должно вызывать в постсинаптической клетке абсолютно такие же физиологические эффекты, что и при раздражении пресинаптического волокна;
2. должно быть доказано, что это вещество выделяется при активации пресинаптического нейрона;
3. действие вещества должно блокироваться теми же агентами, которые подавляют и естественное проведение сигнала.
- это специализированная структура, обеспечивающая межклеточную передачу сигналов электрической и (или) химической природы.
С помощью синапсов передается информация от рецепторных клеток на дендриты чувствительных нейронов, с одной на другую, с нервной клетки на волокно скелетной мышцы, железистые и другие эффекторные клетки. Через синапсы могут оказываться возбуждающие или тормозные влияния на клетки, активироваться или подавляться их метаболизм и другие функции.
Термин «синапс» ввел И. Шеррингтон в 1897 г. В настоящее время синапсами называют специализированные функциональные контакты между возбудимыми клетками (нервными, мышечными, секреторными), служащие для передачи и преобразования нервных импульсов.
Строение синапса
Электронно-микроскопические исследования выявили, что синапсы имеют три основных элемента: пресинаптическую мембрану, постсинаптическую мембрану и синаптическую щель (рис. 1).
Передача информации через синапс может осуществляться химическим или электрическим путем. Смешанные синапсы сочетают химические и электрические механизмы передачи.

Рис. 1. Основные элементы синапса
Виды синапсов
По механизму передачи возбуждения синапсы подразделяют на электрические и химические.
Электрические синапсы образуются между клетками, формирующими между мембранами плотные щелевые контакты. Ширина щели составляет около 3 нм, и между контактирующими мембранами образуются общие ионные каналы с диаметром поры около 1-2 нм. Через эти каналы и осуществляется передача информации с помощью электрических ионных токов. Через каналы электрических синапсов клетки могут обмениваться также небольшими по размеру сигнальными молекулами органической природы. Названные вещества способны перемещаться в электрических синапсах с большой скоростью в обоих направлениях, и переносимая с их помощью информация также может передаваться в обоих направлениях (в отличие от химических синапсов).
Электрические синапсы имеются уже в эмбриональном мозге и остаются наряду с химическими синапсами в зрелой позвоночных.
Ионные токи, перемещающиеся из пресинаптического нейрона в постсинаптический, вызывают на его мембране колебания разности потенциалов — постинаптический потенциал амплитудой около 1 мВ и могут вызвать генерацию на ней ПД. В свою очередь возникший ПД может вызвать обратный ток ионов через каналы щелевых контактов к пресинаптическому нейрону и становится источником модуляции разности потенциалов на его мембране. Нейрон может формировать щелевые контакты (электрические синапсы) с рядом других нейронов, поэтому практически одновременное протекание ионных токов между ними способствует синхронизации активности группы нервных клеток, связанных этими синапсами. Электрические синапсы чаще выявляются в областях мозга, в которых регистрируется высоко синхронизированная нейронная активность.
Как уже упоминалось ранее, ионные каналы щелевых контактов имеются не только между нервными, но и между глиальными клетками, между гладкими миоцитами, между кардиомиоцитами, между железистыми клетками.
Химические синапсы образуются специализированными структурами двух клеток в области их контакта (рис. 2). Одной из этих клеток, которую называют пресинаптической, обычно является нервная клетка, но ею может быть и специализированная чувствительная клетка иной природы (например, сенсоэпителиальная слуховая или вкусовая клетка, гломусные клетки аортального тельца). Пресинаптическая нервная клетка обычно формирует синапс на другой клетке с помощью мембраны нервного окончания (аксона). В этом случае окончание аксона называют пресинаптической, или аксонной, терминально.
Часть мембраны окончания, обращенную в сторону постсинаптической клетки, называют пресинаптической . Клетку, на которой формируется синаптический контакт, называют постсинаптической , а часть плазматической мембраны клетки, обращенную к пресинаптической мембране, — постсинаптической .
Узкое щелевидное пространство, разделяющее пресинаптическую и постсинаптическую мембраны, называют синаптической щелью (см. рис. 2.). Таким образом, для химических синапсов общими структурными элементами являются пресинаптическая часть (нервное окончание и пресинаптическая мембрана), синаптическая щель, постсинаптическая часть (постсинаптическая мембрана).

Рис. 2. Строение синапса и процессы, осуществляемые в ходе синаптической передачи сигнала
Химические синапсы могут образовываться между двумя нервными клетками с участием отростков и тела клетки. В зависимости от структур нейронов, образующих синаптическое соединение, синапсы делят на аксосоматические, аксоаксональные, аксодендритные, дендродендритные. Синапсы, располагающиеся в пределах ЦНС, называют центральными, а находящиеся вне ЦНС — периферическими. Периферические синапсы передают сигналы нервных волокон на эффекторные органы (мышечные волокна, железистые клетки).
Химические синапсы
Химический синапс - межклеточное образование, которое обеспечивает передачу сигнала с помощью химического посредника-медиатора.
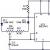
Передача информации в химических синапсах осуществляется через синаптическую щель — область внеклеточного пространства шириной 10-50 нм, разделяющую пре- и постсинаптические мембраны клеток. В пресинаптическом окончании содержатся синаптические везикулы (рис. 3) — мембранные пузырьки диаметром около 50 нм, в каждом из которых заключено 1 . 10 4 — 5 . 10 4 молекул медиатора. Общее количество таких пузырьков в пресинаптических окончаниях составляет несколько тысяч. Цитоплазма синаптической бляшки содержит митохондрии, гладкий эндоплазматический ретикулум, микрофиламенты.
Синаптическая щель заполнена мукополисахаридом, «склеивающим» пре- и постсинаптическую мембраны.

Постсинаптическая мембрана содержит крупные белковые молекулы, выполняющие функции рецепторов, чувствительных к медиатору, а также многочисленные каналы и поры, через которые в пост- синаптический нейрон могут поступать ионы.

Рис. 3. Строение химического синапса
Характеристика химического синапса
- Принцип «физиологического клапана»
- При участии посредника-медиатора
- Синаптическая задержка
- Принцип Дейла
- Трансформация ритма возбуждения
- Синаптическое облегчение и депрессия
- Утомляемость
- Явление суммации, подчинение закону силы
- Низкая лабильность
- Чувствительность к химическим факторам
Передача информации в химических синапсах
При поступлении потенциала действия к пресинаптическому окончанию происходит деполяризация пресинаптической мембраны и повышается ее проницаемость для ионов Са 2+ (рис. 4). Повышение концентрации ионов Са 2+ в цитоплазме синаптической бляшки инициирует экзоцитоз везикул, наполненных медиатором.
Содержимое везикул высвобождается в синаптическую щель, и часть молекул медиатора диффундирует, связываясь с рецептор- ными молекулами постсинаптической мембраны. В среднем каждая везикула содержит около 3000 молекул медиатора, а диффузия медиатора до постсинаптической мембраны занимает около 0,5 мс.
При связывании молекул медиатора с рецептором его конфигурация изменяется, что приводит к открытию ионных каналов и поступлению через постсинаптическую мембрану в клетку ионов, вызывающих развитие потенциала концевой пластинки (ПКП).

Рис. 4. Последовательность событий, происходящих в химическом синапсе от момента возбуждения пресинаптического окончания до возникновения ПД в постсинаптической мембране
ПКП возникает в нервно-мышечных синапсах, в остальных — возбуждающий постсинаптический потенциал (ВПСП) или тормозной постсинаптический потенциал (ТПСП). ПКП есть результат местного изменения проницаемости постсинаптической мембраны для ионов Na + , К + и СI. ПКП не активирует другие хемовозбудимые каналы постсинаптической мембраны, и его величина зависит от концентрации медиатора, действующего на мембрану: чем больше концентрация медиатора, тем выше (до определенного предела) ПКП (ВПСП и ТПСП). Таким образом, ПКП (ВПСП, ТПСП), в отличие от потенциала действия, градуален. При достижении ПКП (ВПСП) некоторой пороговой величины возникают местные токи между участком деполяризованной постсинаптической мембраны с соседними участками электровозбудимой мембраны, что вызывает генерацию потенциала действия.
Если медиатор вызывает открытие Na + -каналов, то возникает ВПСП (по типу деполяризации); если медиатор открывает К+ и СI- каналы, то развивается ТПСП (по типу гиперполяризационного торможения).
Таким образом, процесс передачи возбуждения через химический синапс может быть схематически представлен в виде следующей цепи явлений: потенциал действия на пресинаптической мембране → поступление ионов Ca 2 i внутрь нервного окончания → освобождение медиатора → диффузия медиатора через синаптическую щель к постсинаптической мембране → взаимодействие медиатора с рецептором → активация хемовозбудимых каналов постсинаптической мембраны возникновение потенциала концевой пластинки (ВПСП) критическая деполяризация постсинаптической электровозбудимой мембраны → генерация потенциала действия.
Медиаторы - это биологически активные вещества, посредством которых осуществляются межклеточные взаимодействия в синапсах. К ним относятся ацетилхолин, катехоламины: адреналин, норадре- налин, дофамин; серотонин, гистамин, простагландины, глицин, гамма-аминомасляная кислота (ГАМК). ГАМК и глицин — наиболее распространенные медиаторы синаптического торможения.
В 1935 г. Г. Дейлом было сформулировано правило (принцип Дейла), согласно которому каждая нервная клетка выделяет только один определенный медиатор. Поэтому принято обозначать нейроны по типу медиатора, который выделяется в их окончаниях. Так, нейроны, освобождающие ацетилхолин, называются холинергическими, норадреналин — адренергическими, серотонин — серотонинерги- ческими, амины — аминергическими и т.д.
Химические синапсы имеют два общих свойства:
- возбуждение через химический синапс передается только в одном направлении — от пресинаптической мембраны к постсинаптической мембране (одностороннее проведение);
- возбуждение проводится через синапс значительно медленнее, чем по нервному волокну (синаптическая задержка).
Односторонность проведения обусловлена высвобождением медиатора из пресинаптической мембраны и локализацией рецепторов на постсинаптической мембране. Замедление проведения через синапс (синаптическая задержка) возникает вследствие того, что проведение является многоэтапным процессом (секреция медиатора, диффузия медиатора к постсинаптической мембране, активация хеморецепторов, рост ПКП до пороговой величины) и для протекания каждого из перечисленных этапов требуется время. Кроме того, наличие относительно широкой синаптической щели препятствует проведению импульса с помощью локальных токов.
Особенности строения и функционирования электрических синапсов
Электрический синапс — межклеточное образование, которое обеспечивает передачу импульса возбуждения посредством возникновения электрического тока между пресинаптическим и постсинаптическим отделами.
Электрические синапсы широко распространены в нервной системе беспозвоночных, а у млекопитающих встречаются крайне редко. Вместе с тем электрические синапсы у высших животных широко распространены в сердечной мышце, гладкой мускулатуре, в печени, эпителиальной и железистых тканях.
Ширина синаптической щели в электрических синапсах составляет всего 2-4 нм, что значительно меньше, чем в химических синапсах. Важной особенностью электрических синапсов является наличие между пре- и постсинаптической мембранами своеобразных мостиков, образованных белковыми молекулами, - нексусов. Они представляют собой каналы шириной 1-2 нм (рис. 5).
Свойства электрических синапсов
- Быстродействие (значительно превосходит в химических синапсах)
- Слабость следовых эффектов (практически отсутствует суммация последовательных сигналов)
- Высокая надежность передачи возбуждения
- Пластичность
- Одно- и двухсторонность передачи

Рис. 5. Структура электрического синапса. Характерные особенности: узкая (2-4 нм) синаптическая щель и наличие каналов, образованных белковыми молекулами
Благодаря наличию каналов, размеры которых позволяют переходить из клетки в клетку неорганическим ионам и даже небольшим молекулам, электрическое сопротивление такого синапса, получившего название щелевого или высокопроницаемого контакта, оказывается очень низким. Такие условия позволяют пресинаптическому току распространяться на постсинаптическую клетку практически без угасания.
Электрические синапсы обладают рядом специфических функциональных свойств:
- синаптическая задержка практически отсутствует, т.е. интервал между приходом импульса в пресинаптическое окончание и началом постсинаптического потенциала отсутствует;
- в электрических синапсах двустороннее проведение, хотя стереометрические особенности синапса делают проведение в одном направлении более эффективным;
- электрические синапсы, в отличие от химических, могут обеспечить передачу только одного процесса — возбуждения;
- электрические синапсы менее подвержены воздействию различных факторов (фармакологических, термических и т.д.).
Наряду с химическими и электрическими синапсами, у некоторых нейронов имеются так называемые смешанные синапсы. Их главная особенность заключается в том, что электрическая и химическая передача осуществляется параллельно, поскольку щель между пре- и постсинаптической мембранами имеет участки со структурой химического и электрического синапсов.